A recent study uncovers a trophic cascade after great white sharks disappear
Great white sharks are known to be top-level predators in the marine food web. But what happens when we lose that predator from a marine ecosystem? An ongoing case study in South Africa is highlighted by a recent paper.
Great white sharks have mostly disappeared from South Africa
Apex predators famously regulate the populations of species lower down in the food chain (i.e., mesopredators). They do this both directly by eating these species as prey, and indirectly by altering behaviour and distribution of those same species (Wallach et al. 2015). As such, given that apex predators are among the most threatened globally (Ripple et al. 2014), scientists are eager to understand the effects of losing such a predator on its ecosystem. One regularly studied effect is that of the top-down trophic cascade: in which the loss of the apex predator at the top of the food web has a ripple effect on lower levels (Baum & Worm 2009; Estes et al. 2011).
Although sharks play a wide range of ecological roles, large species such as the great white shark (Carcharodon carcharias) are often regarded as apex predators (Dedman et al. 2024). Indeed, one classic case study of a proposed trophic cascade is that of the loss of large sharks. In 2007, a study in Science (Myers et al. 2007) concluded that the fishing-induced declines of 11 “apex predatory” sharks on the east coast of the United States caused population explosions of their prey such as crowned rays (Figure 1). This population expansion then supposedly led to populations of the rays’ own prey, bay scallops, being decimated, reducing ecosystem productivity and stability, and even causing the closure of local scallop fisheries (Myers et al. 2007). This kind of ripple effect is a textbook trophic cascade (Baum & Worm 2009; Estes et al. 2011); yet its occurrence following the loss of top-predator sharks has come under scrutiny after this nearly-20-year-old study’s publication. A more recent study has argued that there was little correlation between the loss of the sharks and the recorded increase in crowned ray abundance and decline of bay scallops, due to the slow-growing rays also being affected by fishing pressure and a lack of consideration for other bay scallop predators (Grubbs et al. 2016). Furthermore, empirical studies focusing on coral reef habitats have since found that fishing pressure resulted in simultaneous declines across all shark trophic levels: in other words, both apex predators and mesopredators were lost (Roff et al. 2016, Desbiens et al. 2021). Despite this, there is at least one other study supporting evidence for mesopredator population increases following declines of apex predatory sharks (Ferretti et al. 2010). Moreover, the fossil record may show evidence for a global trophic cascade following the extinction of the largest shark of all, Otodus megalodon. Namely, following the giant shark’s extinction in the Pliocene, fossil cetaceans appear to reach larger body sizes than their ancestors after being released from megalodon’s predatory pressure (Pyenson & Sponberg 2011; Pimiento & Clements 2014; Cooper et al. 2022). When all this literature is considered together, the evidence for a trophic cascading effect in the case of losing apex predatory sharks seems mixed at best.
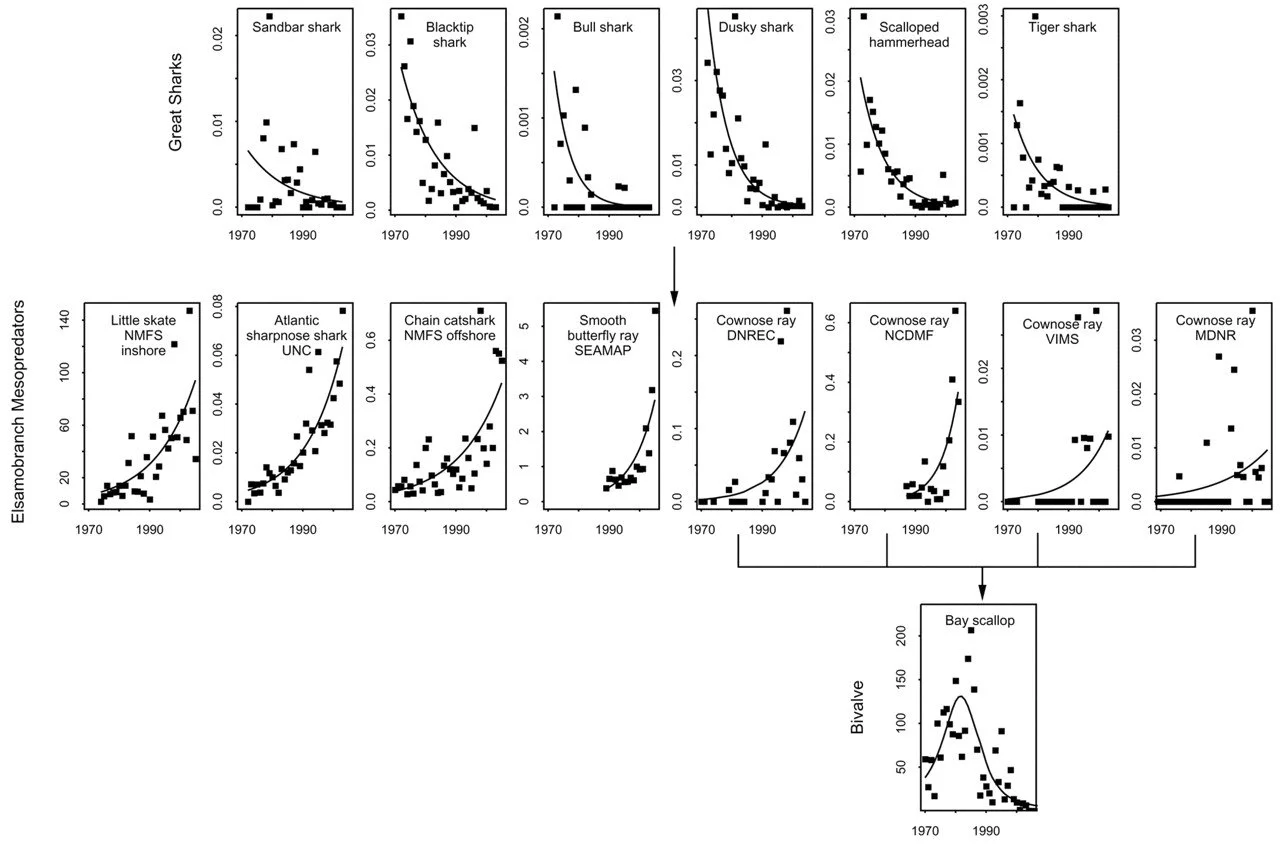
Figure 1. The classic case study of a trophic cascade when apex predatory sharks are lost. This shows change in relative abundance over time in species at each trophic level as estimated from individual data sources: apex predatory sharks (top), elasmobranch mesopredators like the crowned rays (middle), and bay scallops (bottom). This figure is sourced from Figure 1 of Myers et al. (2007).
Yet a strange and worrying happening in South Africa’s Western Cape has provided a unique opportunity to identify such an effect as it happens. Despite being a known hotspot for great white sharks for decades (Cliff et al. 1989; Towner et al. 2013; Bowlby et al. 2023), Western Cape populations of this iconic species (Figure 2) have recently plummeted, leading to outright disappearances in the sites of False Bay, Gansbaai and Mossel Bay (Micarelli et al. 2021; Fisher 2021; Bowlby et al. 2022; 2023; but see also Gennari et al. 2024; Bowlby et al. 2024).

Figure 2. Great white shark aggregation sites in South Africa, marked in solid circles. The three leftmost sites from the Western Cape – False Bay, Gansbaai and Mossel Bay – are the sites where great white sharks have declined in recent years. This figure is sourced from Figure 1 of Bowlby et al. (2023).
On the surface, one cause seems clear in Gansbaai and Mossel Bay: antipredator responses to the presence of killer whales (Orcinus orca). Since 2017, white sharks in Gansbaai, and later Mossel Bay, have been killed by killer whales for their livers; most famously by a pair of males with collapsed dorsal fins known as Port and Starboard (Figure 3; Towner et al. 2022; 2023; 2024).

Figure 3. Port (left) and Starboard (right), two male killer whales that are known predators of great white sharks in South Africa. Photo taken by Esther Jacobs.
I have a personal history with this story, having witnessed the necropsy of the fifth known victim of Port and Starboard, a 4.1 m male (Figure 4; Towner et al. 2022), while I was an intern at White Shark Diving Company in Gansbaai. As such, I’ve seen some of the fallout of these killings first hand, and have recently written about them in an article for the Guardian (Cooper 2025). To sum it up, although white sharks have been hunted by killer whales in California (Pyle et al. 1999) and most recently in Australia (Reeves et al. 2025); the effects seen in South Africa have been the most dramatic. As I detailed in my Guardian article (Cooper 2025), the white sharks have all but disappeared from Gansbaai and Mossel Bay, vanishing for longer and longer periods of time as the killer whales have struck, and returning in dwindling numbers.

Figure 4. The necropsy of a 4.1 m male great white shark that was killed by killer whales for its liver, recovered on June 24th 2017 (Towner et al. 2022). The killers were most likely Port and Starboard, who were seen in the area at around the same time the necropsy was being conducted. Included as part of the necropsy are Mary Rowlinson (left), who headed my internship at White Shark Diving Company, and Dr Alison Towner (middle), who led the necropsy and all subsequent studies on Port and Starboard (Towner et al. 2022; 2023; 2024). Photo taken by Jack Cooper.
By stark contrast, no such killings of white sharks have been detected in False Bay; yet the white shark population there began to decline in 2015 and has disappeared altogether since as long ago as 2018 (Hammerschlag et al. 2019; 2025). Such effects already documented include the emergence of the sevengill shark (Notorynchus cepedianus) to the area, seemingly taking the white sharks’ place as the local “apex predator” following their disappearance (Hammerschlag et al. 2019). Given this drastic change, a new paper (Hammerschlag et al. 2025) has taken the opportunity to investigate if a trophic cascade may have occurred in False Bay.
The new study
Three different methods were used in this new study, focusing entirely on the local ecosystem in False Bay (Hammerschlag et al. 2025). To estimate the abundance of both white sharks and sevengill sharks, the authors conducted standardised boat-based surveys between the years 2000 and 2020. Sharks were attracted using decoy seal baits and baited lines, with individuals being identified based on unique markers such as scarring. Relative abundance was calculated using the observed sharks per hour, which was then converted into annual means and mean values in the period before (2000-2015; “pre-loss”) and after (2016-2020; “post-loss”) the white sharks disappeared from False Bay. The change in white shark abundance was drastic: going from an average of 1.64 sharks an hour in 2000-2015 to just 0.3 sharks an hour in 2016-2020, marking an 82% decline (Figure 5). Indeed, not a single white shark was sighted in False Bay after August 2018. By contrast, no sevengill sharks were seen at all while white sharks were abundant but rose to 0.96 sharks an hour in 2016-2020 when the white sharks declined and finally disappeared (Figure 6). As such, this supports previous work suggesting that sevengill sharks, which can be prey of white sharks, emerged to fill in the newly vacant trophic niche (Hammerschlag et al. 2019).
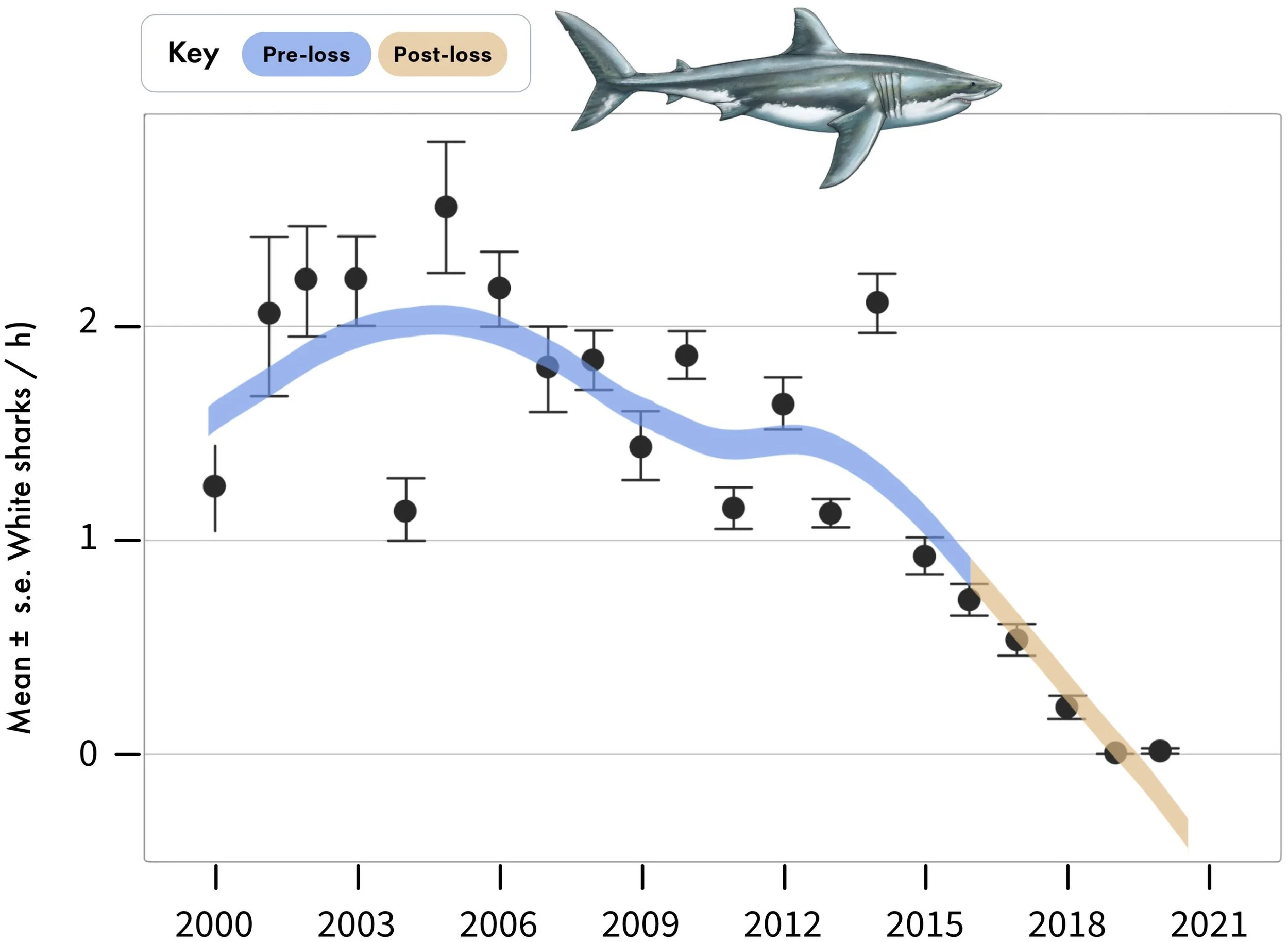
Figure 5. Annual mean and standard error sightings of the great white shark in False Bay between 2000 and 2020. This figure is sourced from Figure 2 of Hammerschlag et al. (2025). Pre- and post-loss refer to periods prior to and following the decline of great white sharks respectively.

Figure 6. Annual mean and standard error sightings of the sevengill shark in False Bay between 2000 and 2020. This figure is sourced from Figure 6 of Hammerschlag et al. (2025). Pre- and post-loss refer to periods prior to and following the decline of great white sharks respectively.
Secondly, the team used publicly available data from iNaturalist and the Global Biodiversity Information Facility (GBIF) to analyse False Bay sightings of cape fur seals (Arctocephalus pusillus), animals that white sharks are well known to prey on in South Africa, including in False Bay (Martin et al. 2005; Hammerschlag et al. 2006; Martin & Hammerschlag 2012). What they found was that cape fur seal observations in False Bay increased by a whopping 520% once the white sharks disappeared. Moreover, their geographical range in False Bay grew by an enormous 846% between 2016 and 2019 right as the white sharks vanished (Figure 7; Hammerschlag et al. 2025). The clear correlation between these increases and the disappearance of the white sharks points towards a predator release effect, a telltale sign of a trophic cascading effect (Myers et al. 2007).

Figure 7. Annual sightings of the cape fur seal in False Bay between 2000 and 2020. This figure is sourced from the top half of Figure 5 of Hammerschlag et al. (2025). Pre- and post-loss refer to periods prior to and following the decline of great white sharks respectively.
Finally, baited remote underwater stations (BRUVs) were deployed within False Bay, with footage analysed for species count as another metric of relative abundance of sharks. Despite only three white sharks being detected in 2012 likely representing an under-sample, none were spotted in the “post-loss” period (2016-2020). Meanwhile, sevengill sharks were sighted only in the “post-loss” period, mirroring the boat survey results. What the BRUVs data added, however, was the abundance of cape horse mackerel (Trachurus capensis), which are known prey of the cape fur seals (Hammerschlag et al. 2025). The data found, following the disappearance of the white sharks, mackerel abundance declined by 22% during summer, typically their most abundant season. Similar seasonal declines of 72% and 42% respectively were also detected for pyjama sharks (Poroderma africanum) and common smoothhound sharks (Mustelus mustelus), known prey of sevengill sharks (Hammerschlag et al. 2025). These findings occurring alongside the huge expansion of the cape fur seal population and the emergence of sevengill sharks is likely no coincidence. Collectively, these results all showcase what appears to be trophic cascading effects following the loss of the white sharks – with cape fur seals and sevengill sharks expanding their numbers while their prey subsequently crash (Figure 8).
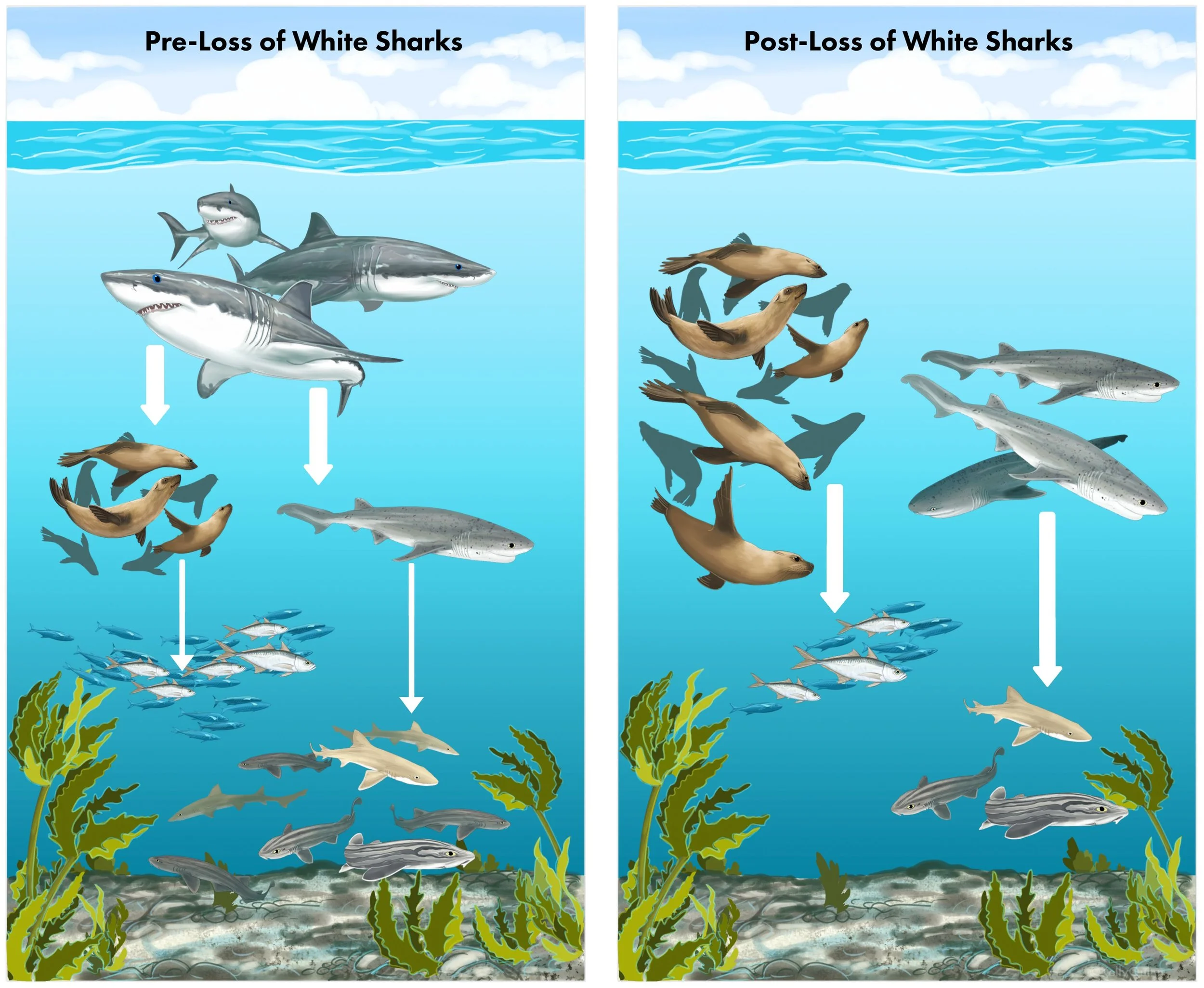
Figure 8. The graphical abstract from Hammerschlag et al. (2025) that summarises the key result of the study. The left panel denotes the food web before the disappearance of great white sharks, while the right panel represents the resulting food web following the disappearance, showcasing the increased abundance of cape fur seals and sevengill sharks and the declines of lower-level prey.
The wider implications for South Africa and shark conservation
With evidence for a trophic cascade in False Bay shown in the new paper (Hammerschlag et al. 2025), it makes me think about similar effects we might be seeing elsewhere in South Africa. After all, with white sharks disappearing following killer whale predations, including by the infamous Port and Starboard (Figure 3), in Gansbaai and Mossel Bay (Towner et al. 2022; 2023; 2024), it’s worth looking at these sites to see what effects have been had. Indeed, I briefly discussed this with Dr Alison Towner and Esther Jacobs in my article for the Guardian (Cooper 2025). In Gansbaai, the white shark cage diving industries have had to rely on bronze whaler sharks (Carcharhinus brachyurus) as their main attraction in the aftermath of the killer whale killings (A. Towner, Pers. Comm). Indeed, these sharks, another known prey of white sharks, have dramatically increased in abundance since the white sharks started declining there (Fisher 2021), just like the sevengill sharks in False Bay (Hammerschlag et al. 2019; 2025). While no data has yet been published to my knowledge for Mossel Bay (E. Jacobs, Pers. Comm.); cape fur seal populations in both Gansbaai and Mossel Bay have also expanded (Fisher 2021). Indeed, cape fur seals in Mossel Bay have recently tested positive for rabies, and locals there believe that the prevalence of white sharks may have reduced this outbreak since white sharks would have most likely picked off the sick individuals (Cooper 2025). This may indicate that trophic cascades are also occurring in Gansbaai and Mossel Bay and is something that ought to be investigated further by teams on the ground there. In the case of these two sites, it seems that the loss of white sharks can be traced back to the killer whale predations (Towner et al. 2022; 2023; 2024), such as those by Port and Starboard (Figure 3).
Exactly what caused the white shark disappearance in False Bay, however, is still a bit of a mystery. While Port and Starboard have been seen killing the newly arrived sevengill sharks in False Bay (Engelbrecht et al. 2019), they have no documented kills of white sharks in the area (Hammerschlag et al. 2025). It is possible that the mere presence of Port and Starboard, who were first documented in South Africa in 2015 (Towner et al. 2022), right as white sharks started leaving False Bay, may have been enough to scare off the white sharks. Indeed, similar anti-predator responses to killer whales have already been documented by white sharks in the northeastern Pacific Ocean (Jorgensen et al. 2019). Whatever the cause of the disappearances in False Bay, the significance of this new study is how it highlights the effects of the loss of the great white shark in its local ecosystems – something that should both alarm us and get us thinking about what might happen if we lose the white shark, and indeed any large predatory shark species, worldwide.
Today, over a third of all shark species are threatened with extinction; their global populations having halved since 1970 (Dulvy et al. 2014; 2021; 2024). The great white shark is among those threatened species, listed as Vulnerable by the IUCN Red List (Rigby et al. 2022). The overwhelming leading threat towards sharks is overfishing (Dulvy et al. 2014; 2021; 2024), something that the white shark is vulnerable to given its low fecundity (i.e., litter size) and slow growth rate (Bruce 2008; Fisher 2021; Bowlby et al. 2022; O’Connell et al. 2024). The effects of their loss in South Africa (Towner et al. 2022; Cooper 2025; Hammerschlag et al. 2025) should be a warning to us about what we might see in the coming decades if we do not regulate worldwide fisheries driving extinction risk and decline of sharks in every ocean.
Acknowledgements and further reading
I thank the staff of White Shark Diving Company for allowing me to observe the white shark dissection in Gansbaai; as well as them and the staff of White Shark Africa for having me as an intern in 2017 and 2018. Further, I thank Dr Alison Towner and Esther Jacobs for contributing their expertise to my original Guardian article, which detailed similar trophic cascades in Gansbaai and Mossel Bay to what has been described in False Bay by Hammerschlag et al. 2025.
You can read the full paper at Frontiers in Marine Science under the following reference:
Hammerschlag N, Herskowitz Y, Fallows C & Couto T, 2025. Evidence of cascading ecosystem effects following the loss of white sharks from False Bay, South Africa. Frontiers in Marine Science, 12, 1530362.
You can also read my Guardian article on the killer whale killings of white sharks in Gansbaai and Mossel Bay at: https://www.theguardian.com/environment/2025/jan/23/south-africa-gansbaai-marine-biology-apex-predators-orcas-conquer-great-white-sharks-seals-penguins-trophic-cascade-aoe.
References
Baum JK & Worm B, 2009. Cascading top‐down effects of changing oceanic predator abundances. Journal of Animal Ecology, 78, 699-714.
Bowlby HD, Dicken ML, Towner AV, Waries S, Rogers T & Kock A, 2023. Decline or shifting distribution? A first regional trend assessment for white sharks (Carcharodon carcharias) in South Africa. Ecological Indicators, 154, 110720.
Bowlby HD, Dicken ML, Towner AV, Rogers T, Waries S & Kock A, 2024. Ecological conclusions remain unchanged for white sharks in South Africa: A reply to Gennari et al. 2024. Ecological Indicators, 165, 112160.
Bowlby HD, Hammerschlag N, Irion DT & Gennari E, 2022. How continuing mortality affects recovery potential for prohibited sharks: The case of white sharks in South Africa. Frontiers in Conservation Science, 3, 988693.
Bruce BD, 2008. The biology and ecology of the white shark, Carcharodon carcharias. In Sharks of the open ocean: Biology, fisheries and conservation (pp. 69–81). Oxford: Blackwell Publishing.
Cliff G, Dudley SFJ & Davis B, 1989. Sharks caught in the protective gill nets off Natal, South Africa. 2. The great white shark Carcharodon carcharias (Linnaeus). South African Journal of Marine Science, 8, 131-144.
Cooper JA, 2025. ‘Awe-inspiring and harrowing’: how two orcas with a taste for liver decimated the great white shark capital of the world. The Guardian. https://www.theguardian.com/environment/2025/jan/23/south-africa-gansbaai-marine-biology-apex-predators-orcas-conquer-great-white-sharks-seals-penguins-trophic-cascade-aoe.
Cooper JA, Hutchinson JR, Bernvi DC, Cliff G, Wilson RP, Dicken ML, Menzel J, Wroe S, Pirlo J & Pimiento C, 2022. The extinct shark Otodus megalodon was a transoceanic superpredator: Inferences from 3D modeling. Science Advances, 8, eabm9424.
Dedman S, Moxley JH, Papastamatiou YP, Braccini M, Caselle JE, Chapman DD, Cinner JE, Dillon EM, Dulvy NK, Dunn RE, Espinoza M, Harborne AR, Harvey ES, Heupel MR, Huveneers C, Graham NAJ, Ketchum JT, Klinard NV, Kock AA, Lowe CG, MacNeil MA, Madin EMP, McCauley DJ, Meekan MG, Meier AC, Simpfendorfer CA, Tinker MT, Winton M, Wirsing AJ & Heithaus MR, 2024. Ecological roles and importance of sharks in the Anthropocene Ocean. Science, 385, adl2362.
Desbiens AA, Roff G, Robbins WD, Taylor BM, Castro‐Sanguino C, Dempsey A & Mumby PJ, 2021. Revisiting the paradigm of shark‐driven trophic cascades in coral reef ecosystems. Ecology, 102, e03303.
Dulvy NK, Fowler SL, Musick JA, Cavanagh RD, Kyne PM, Harrison LR, Carlson JK, Davidson LN, Fordham SV, Francis MP, Pollock CM, Simpfendorfer CA, Burgess GH, Carpenter KE, Compagno LJV, Ebert DA, Gibson C, Heupel MR, Livingstone SR, Sanciangco JC, Stevens JD, Valenti S, & White WT, 2014. Extinction risk and conservation of the world's sharks and rays. Elife, 3, e00590.
Dulvy NK, Pacoureau N, Matsushiba JH, Yan HF, VanderWright WJ, Rigby CL, Finucci B, Sherman CS, Jabado RW, Carlson JK, Pollom RA, Charvet P, Pollock CM, Hilton-Taylor C & Simpfendorfer CA, 2024. Ecological erosion and expanding extinction risk of sharks and rays. Science, 386, eadn1477.
Dulvy NK, Pacoureau N, Rigby CL, Pollom RA, Jabado RW, Ebert DA, Finucci B, Pollock CM, Cheok J, Derrick DH, Herman KB, Sherman CS, VanderWright WJ, Lawson JM, Walls RHL, Carlson JK, Charvet P, Bineesh KK, Fernando D, Ralph GM, Matsushiba JH, Hilton-Taylor C, Fordham SV & Simpfendorfer CA, 2021. Overfishing drives over one-third of all sharks and rays toward a global extinction crisis. Current Biology, 31, 4773-4787.
Engelbrecht TM, Kock AA & O'Riain MJ, 2019. Running scared: when predators become prey. Ecosphere, 10, e02531.
Estes JA, Terborgh J, Brashares JS, Power ME, Berger J, Bond WJ, Carpenter SR, Essington TE, Holt RD, Jackson JB, Marquis RJ, Oksanen L, Oksanen T, Paine RT, Pikitch EK, Ripple WJ, Sandin SA, Scheffer M, Schoener TW, Shurin JB, Sinclair AR, Soule ME, Virtanen R & Wardle DA, 2011. Trophic downgrading of planet Earth. Science, 333, 301-306.
Ferretti F, Worm B, Britten GL, Heithaus MR & Lotze HK, 2010. Patterns and ecosystem consequences of shark declines in the ocean. Ecology Letters, 13, 1055-1071.
Fisher R, 2021. Possible causes of a substantial decline in sightings in South Africa of an ecologically important apex predator, the white shark. South African Journal of Science, 117, 1-4.
Gennari E, Hammerschlag N, Andreotti S, Fallows C, Fallows M & Braccini M, 2024. Uncertainty remains for white sharks in South Africa, as population stability and redistribution cannot be concluded by Bowlby et al. (2023):“Decline or shifting distribution? a first regional trend assessment for white sharks (Carcharodon carcharias) in South Africa. Ecological Indicators, 160, 111810.
Grubbs RD, Carlson JK, Romine JG, Curtis TH, McElroy WD, McCandless CT, Cotton CF & Musick JA, 2016. Critical assessment and ramifications of a purported marine trophic cascade. Scientific Reports, 6, 20970.
Hammerschlag N, Herskowitz Y, Fallows C & Couto T, 2025. Evidence of cascading ecosystem effects following the loss of white sharks from False Bay, South Africa. Frontiers in Marine Science, 12, 1530362.
Hammerschlag N, Martin RA & Fallows C, 2006. Effects of environmental conditions on predator–prey interactions between white sharks (Carcharodon carcharias) and Cape fur seals (Arctocephalus pusillus pusillus) at Seal Island, South Africa. Environmental Biology of Fishes, 76, 341-350.
Hammerschlag N, Williams L, Fallows M & Fallows C, 2019. Disappearance of white sharks leads to the novel emergence of an allopatric apex predator, the sevengill shark. Scientific Reports, 9, 1908.
Jorgensen SJ, Anderson S, Ferretti F, Tietz JR, Chapple T, Kanive P, Bradley RW, Moxley JH & Block BA, 2019. Killer whales redistribute white shark foraging pressure on seals. Scientific Reports, 9, 6153.
Martin RA, Hammerschlag N, Collier RS & Fallows C, 2005. Predatory behaviour of white sharks (Carcharodon carcharias) at Seal Island, South Africa. Journal of the Marine Biological Association of the United Kingdom, 85, 1121-1136.
Martin RA & Hammerschlag N, 2012. Marine predator–prey contests: ambush and speed versus vigilance and agility. Marine Biology Research, 8, 90-94.
Micarelli P, Bonsignori D, Compagno LJV, Pacifico A, Romano C & Reinero FR, 2021. Analysis of sightings of white sharks in Gansbaai (South Africa). The European Zoological Journal, 88, 363-374.
Myers RA, Baum JK, Shepherd TD, Powers SP & Peterson CH, 2007. Cascading effects of the loss of apex predatory sharks from a coastal ocean. Science, 315, 1846-1850.
O’Connell CP, Dodd JF, Crews J, Gressle J, Racicot B, Sitzer S, Lis T & Skomal GB, 2024. A global review of white shark (Carcharodon carcharias) parturition. Reviews in Fish Biology and Fisheries, 34, 869-893.
Pimiento C & Clements CF, 2014. When did Carcharocles megalodon become extinct? A new analysis of the fossil record. PLoS One, 9, e111086.
Pyenson ND & Sponberg SN, 2011. Reconstructing body size in extinct crown Cetacea (Neoceti) using allometry, phylogenetic methods and tests from the fossil record. Journal of Mammalian Evolution, 18, 269-288.
Pyle P, Schramm MJ, Keiper C & Anderson SD, 1999. Predation on a white shark (Carcharodon carcharias) by a killer whale (Orcinus orca) and a possible case of competitive displacement. Marine Mammal Science, 15, 563-568.
Reeves IM, Weeks AR, Towner AV, Impey R, Fish JJ, Clark ZS, Butcher PA, Meyer L, Donnelly DM, Huveneers C, Hudson N & Miller AD, 2025. Genetic evidence of killer whale predation on white sharks in Australia. Ecology and Evolution, 15, e70786.
Rigby CL, Barreto R, Carlson J, Fernando D, Fordham S, Francis MP, Herman K, Jabado RW, Jones GCA, Liu KM, Lowe CG, Marshall A, Pacoureau N, Romanov E, Sherley RB & Winker H, 2022. [amended version of 2019 assessment]. “Carcharodon carcharias”. 2022, e.T3855A212629880.
Ripple WJ, Estes JA, Beschta RL, Wilmers CC, Ritchie EG, Hebblewhite M, Berger J, Elmhagen B, Letnic M, Nelson MP, Schmitz OJ, Smith DW, Wallach AD & Wirsing AJ, 2014. Status and ecological effects of the world’s largest carnivores. Science, 343, 1241484.
Roff G, Doropoulos C, Rogers A, Bozec YM, Krueck NC, Aurellado E, Priest M, Birrell C & Mumby PJ, 2016. The ecological role of sharks on coral reefs. Trends in Ecology & Evolution, 31, 395-407.
Towner AV, Kock AA, Stopforth C, Hurwitz D & Elwen SH, 2023. Direct observation of killer whales preying on white sharks and evidence of a flight response. Ecology, 104, e3875.
Towner A, Micarelli P, Hurwitz D, Smale MJ, Booth AJ, Stopforth C, Jacobs E, Reinero FR, Ricci V, Di Bari A, Gavazzi S, Caurgno G, Mahrer M & Gennari E, 2024. Further insights into killer whales Orcinus orca preying on white sharks Carcharodon carcharias in South Africa. African Journal of Marine Science, 46, 1-5.
Towner AV, Watson RGA, Kock AA, Papastamatiou Y, Sturup M, Gennari E, Baker K, Booth T, Dicken M, Chivell W, Elwen S, Kaschke T, Edwards D & Smale M, 2022. Fear at the top: killer whale predation drives white shark absence at South Africa’s largest aggregation site. African Journal of Marine Science, 44, 139-152.
Towner AV, Wcisel MA, Reisinger RR, Edwards D & Jewell OJ, 2013. Gauging the threat: the first population estimate for white sharks in South Africa using photo identification and automated software. PLoS One, 8, e66035.
Wallach AD, Izhaki I, Toms JD, Ripple WJ & Shanas U, 2015. What is an apex predator? Oikos, 124, 1453-1461.