A tribute to Dr Gordon Hubbell and his academic works
I was saddened to hear of the passing of Dr Gordon Hubbell on June 14th 2026. Like many in the field of shark palaeobiology, both academic and enthusiast, I greatly admired Gordon’s wonderful collection of shark fossils – the best in the world in my opinion. He has also contributed a number of academic works that made significant leaps in our knowledge of sharks. So, I thought I’d write this piece to share my own personal memories of Dr Hubbell and reflect on his academic works and contributions.
A memory of Dr Hubbell
June 27th 2022
It is a hot, humid day in Gainesville, Florida, and I am sweating so much that my face is practically a waterfall. I’ve been drinking Gatorade – a drink I’d never even touched before last week – virtually non-stop just to stay hydrated. But the heat is not the only reason I’m breaking a sweat. My destination is the other.
I’ve come to Gainesville to collect crucial tooth measurement data from Cenozoic shark teeth in the collections of the University of Florida; with the goal of informing two of my three chapters of my PhD. Specifically, I want to first understand if these tooth measurements can inform shark functional traits (published in Cooper et al. 2023) and then use this information to understand how shark functional diversity changed over the last 66 million years (published in Cooper & Pimiento 2024). Right now, I have the day off from the Florida Museum collection and am sitting in an uber heading for the private, but accessible, collection of Dr Gordon Hubbell.
Gordon Hubbell is a man I’ve admired from afar for years. Why? Because he happens to be a world-leading authority on fossil sharks. He had retired to Gainesville after a career in veterinary medicine, and has built the most impressive collection of fossil and living shark specimens anywhere on Earth (Figure 1). I’d first come across Gordon’s collection on a National Geographic documentary at around 10 years old, a few years after my interest in megalodon had kicked off. I was mind blown at how one man had such passion that he had essentially built a museum in his own house. The curator of the Florida Museum, Bruce MacFadden, worked closely with him and produced multiple PhD graduates who used Gordon’s material in their study of sharks and continue to study fossil sharks to this day. Among them were Dr Dana Ehret, Dr Victor Perez, and Dr Catalina Pimiento, who would go on to become my own PhD supervisor. Most excitingly for me was the fact that Gordon’s collection contained the largest megalodon tooth currently on scientific record (GHC 6; a tooth described in Perez et al. 2021; Figure 2).

Figure 1. Gordon stands within his collection for a press photo. A wall of shark jaws shows just how wide the range of species was in his collection, even of living species. Photo by Bethany Gaffey; sourced from: https://www.themarinediaries.com/tmd-blog/dr-gordon-hubbell-bitten-by-the-shark-tooth-bug.

Figure 2. Specimen “GHC-6” (Perez et al. 2021): the largest megalodon tooth currently documented in the scientific record. Photo taken by myself during my visit to Gordon’s collection in June 2022; and used as data within Cooper & Pimiento (2024).
Now, as I get out the Uber and stand in front of Dr Hubbell’s house, I feel a huge smile on my face knowing that I’m about to see this collection and spend a few hours in it collecting data. I’d already spoken to Gordon on the phone to arrange today, but now I’m getting to meet him face to face. To me, a lad who’s shared that interest in fossil sharks for most of my life, I might as well be meeting Sir David Attenborough.
Gordon opens the door and greets me with a smile. He’s in his late 80s at the time but still has the eager eye of an excited youngling. He invites me in and we spend the next three hours photographing and measuring all the specimens needed for my data. I see the huge megalodon tooth (Figure 2), jaws of so many different shark species, the incredible dentition of Carcharodon hubbelli (Figure 3), a megalodon vertebra from Peru, entire Otodus dentitions, and teeth of species only available in this collection. When I mention that I will be travelling on to Peru next, he tells me about several travels of his to Peru, including where he found the megalodon vertebra. His enthusiasm was so infectious that I probably wasted around 30 minutes of my data collecting just chatting to him about his adventures. I had to leave by early afternoon so he could run errands, and of course respected that wish, but he was so generous with his time and his stories that I could have stayed there all day.

Figure 3. Specimen UF 226255, the incredible holotype specimen of Carcharodon hubbelli; as kept in Gordon’s collection. Photo taken by myself during my visit to Gordon’s collection in June 2022.
It was a genuine highlight of my entire PhD project. As if the day hadn’t been incredible enough, it was that same day I learned that a paper of mine (Cooper et al. 2022) had been accepted for publication. I guess Gordon was a good luck charm. When I returned to Scotland, I sent him a handwritten letter to thank him for his hospitality and generosity. He was of a similar generation to my late grandmother, and we had communicated with letters while I was at university, so I figured that was the best way. Bruce, who had looked after me during my stay in Gainesville, would later tell me that he had appreciated it, something that made me feel warm to hear.
The academic contributions of Dr Hubbell
I’ve been reflecting on that wonderful experience for the last several days. Gordon stayed active in aiding researchers the following years to the best of my knowledge, still in correspondence with Bruce in particular. At least that was what I heard when I spoke to various people who had worked with him at the 2025 SVP conference in Birmingham.
On the evening of June 14th, 2026, while scrolling social media, I came across the sad news that Gordon had passed away that morning. Born on February 28th 1935, he was 91 years old. I’ve since seen numerous tributes from people far closer to him than I or any researcher was. Having only met him the one time, it wouldn’t be right for me to write a post detailing his life. Instead, I’d like to pay tribute to his numerous academic contributions to our field in the form of his papers.
It surprised me to learn, upon checking the website Shark References, that Gordon only ever published 10 papers (at least 10 that are recorded on the website). Quantity of papers of course means nothing in terms of impact, which came from Gordon’s extensive knowledge. One of these papers listed is a conference abstract and another is a Spanish article that I was unable to track down. So for this blog, as my own little tribute to Gordon, I thought I’d not only share my memory of meeting him; but would go through the 8 remaining papers one by one. Let’s take a look below:
Paper 1: Paleocene-Pliocene sharks from Cuba
The first study highlighted here is a study from 1996 in which Gordon worked alongside Manuel Iturralde-Vinent and Reinaldo Rojas to comprehensively revise Cuba’s fossil shark and ray record across the Cenozoic era. They examined various museum collections, historical records, new specimens and published records to compile the wide taxonomic range of sharks and rays from the Paleocene, Eocene, Miocene and Pliocene epochs in Cuba. From this, they identified 19 species across 10 genera (e.g., Figure 4), ultimately bringing us a full inventory of Cuban elasmobranchs from this time that was known in 1996 (Iturralde-Vinent et al. 1996).
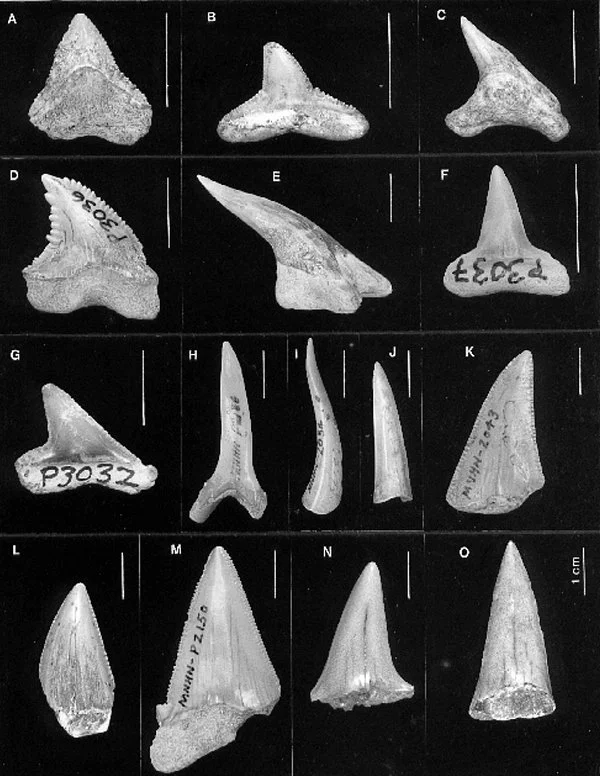
Figure 4. Teeth gathered and figured by Iturralde-Vinent et al. (1996). Details of tooth identification are in the paper, but it should be noted that some of the names are now considered taxonomically invalid. Sourced from Figure 1 of Iturralde-Vinent et al. (1996).
However, what makes this study interesting to me is that it is not simply a catalogue of what sharks swam around Cuba in the Cenozoic. What stands out to me is that it uses these taxa and where they are found geologically as indicators of what ancient Cuban marine environments may have looked like. In this regard, the geographic distribution of Miocene sharks takes particular interest. Specifically, Gordon and his co-authors observed that western Cuba, particularly an area called the Havana-Matanzas region, was dominated by sharks that were large and pelagic – including great white and mako sharks (Carcharodon and Isurus spp; though it should be noted that the taxon identified as Isurus hastalis in this 1996 study is now considered to have been an ancient white shark Carcharodon hastalis; Ehret et al. 2012), and even Otodus megalodon. By comparison, central Cuban localities were filled mainly with sharks that typically live in shallower coastal environments such as lemon sharks (Negaprion brevirostris) and hammerhead sharks (Sphyrna spp).
Gordon and his co-authors ultimately interpreted this sharp difference as evidence of a marine seaway crossing western Cuba and ultimately connecting the Caribbean Sea with the deeper Atlantic Ocean. The catalogued sharks are therefore being used here as essentially palaeoceanographic indicators. The way I like to see it, it shows how different assemblages of shark teeth even in the same broad area can reveal ecological and environmental structures of our ancient oceans. Even these studies that appear simple on paper can reveal a lot of insight into ancient marine ecosystems, and Iturralde-Vinent et al. (1996) certainly fits the bill.
Paper 2: A dental lens into the evolutionary history of the great white shark
This second paper (Hubbell 1996) is a book chapter written by Gordon for the 1996 book Great White Sharks: The Biology of Carcharodon carcharias. This book continues to be a key reference for anybody studying white sharks. One chapter (Gottfried et al. 1996) is even, to this day, one of the keystone studies on megalodon as it was one of the first to “deep dive” into megalodon’s possible anatomy and ecology; even if it is dated now in terms of taxonomy (i.e., megalodon is no longer considered Carcharodon megalodon; see Nyberg et al. 2006; Ehret et al. 2012; Shimada et al. 2017) and anatomy (i.e., see Sternes et al. 2024; Shimada et al. 2025).
Hubbell (1996) is arguably Gordon’s most foundational, particularly with regards to how we link shark teeth from modern sharks to ecological interpretations of the fossil record. It harkens to a key question that continues to partially inform the results of many different papers relying on shark teeth as ecological proxies, including one of my own PhD chapters (Cooper et al. 2023 and references therein): how much variation exists within living white shark teeth and how might that affect our interpretations of the fossil record?
To answer this question, Gordon used 40 modern great white shark jaws from his own collection of individuals ranging from 1.22 to 5.94 m in length. From the teeth, he took six measurements including crown height and crown width (Figure 5). He then normalised all measurements for body size across specimens, allowing the dental characters to be compared across different tooth positions and ontogenetic stage (Hubbell 1996).
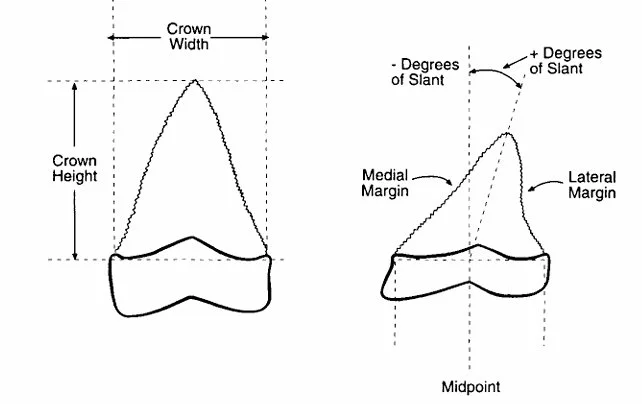
Figure 5. Tooth measurements taken by Gordon as part of his study into white shark teeth. Sourced from Figure 1 of Hubbell (1996).
What he found from this was that the teeth did indeed vary across tooth position and ontogenetic stage. Specifically, great white shark teeth varied rather dramatically in shape, orientation, serration density and proportion. As the shark fossil record is overwhelmingly represented by isolated teeth, this finding had enormous implications. Moreover, adults and juveniles could have very different teeth as well. Adult white shark teeth were broad with refined serrations and no lateral cusplets (Figure 6) whereas juvenile white shark teeth are narrower with small cusplets (Figure 7; Hubbell 1996). Other studies have since found similar results to this (e.g., French et al. 2017) with the juvenile tooth cusplets sometimes being called “serrational cusplets” (Bemis et al. 2015; Cooper et al. 2023). Perhaps most importantly, adult white shark teeth could also have variation with each other – such as in serration counts, broadness and curvature. As such, given that the shark fossil record is dominated overwhelmingly by isolated teeth, Gordon used this study to caution assigning new extinct shark species in the genus Carcharodon or otherwise based on only a small sample of teeth; and that both tooth position and ontogenetic stage should be considered when using fossil teeth as ecological indicators.
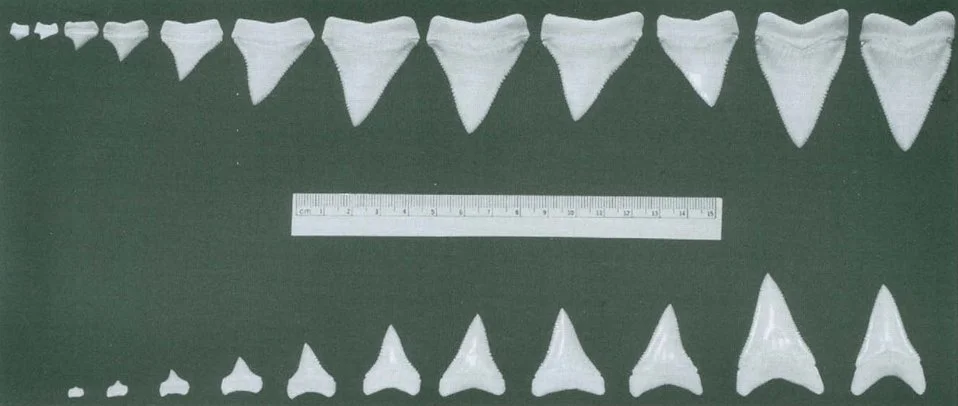
Figure 6. Teeth of a 4.4 m white shark from Gordon’s collection. Sourced from Figure 2 of Hubbell (1996).

Figure 7. Teeth of a 1.42 m white shark from Gordon’s collection. Note the differences in morphology between this juvenile individual and those of the much larger shark from Figure 6: this is a classic case of ontogenetic variation of tooth morphology in sharks. Sourced from Figure 3 of Hubbell (1996).
This study is a particular standout from Gordon as it arguably transformed shark tooth morphology from a purely descriptive tool to a comparative framework that could be used to understand tooth position, ontogeny and, by extension, ecology in extinct sharks. Many, many studies that followed Hubbell (1996) examined white shark evolution, taxonomy and phylogeny based on principles directly from this study. Even my own PhD work needed to consider the role of tooth position when examining how isolated teeth can inform functional traits in sharks (Cooper et al. 2023). Gordon’s paper (Hubbell 1996) ultimately remains one of the clearest demonstrations on how intraspecific variation within shark teeth must be considered before we can even begin to use teeth to reconstruct evolutionary history.
Paper 3: Symmetrical teeth in Cretoxyrhina
A small study Gordon contributed to was a 2001 paper co-authored with Professor Kenshu Shimada (Shimada & Hubbell 2001). Kenshu’s earlier work had focused heavily on the Cretaceous shark Cretoxyrhina mantelli, which at the time was the only extinct lamniform shark with a well-constructed dentition (Shimada 1997a, b). An earlier study (Eastman 1895) had identified two small symmetrical teeth in a C. mantelli tooth set that were considered symphysial. Kenshu’s work, however, had argued that these were instead abnormal teeth (Shimada 1997a). The follow up study with Gordon (Shimada & Hubbell 2001) agreed with this interpretation due to a comparison with one of Gordon’s specimens of Alopias superciliosus, considered phylogenetically close to Cretoxyrhina at the time (Shimada 1999) and which also possessed similarly abnormal teeth across tooth rows – specifically small symmetrical teeth with a remarkable resemblance to those seen in Cretoxyrhina (Shimada & Hubbell 2001).
This paper is an early example of how Gordon’s material was used directly to aid researchers, and one of several papers where Gordon would serve as a co-author in this research. It’s a short but sweet paper that gets straight to the point and may well indicate that dental abnormalities we see in modern sharks also occurred in extinct species from the fossil record (Shimada & Hubbell 2001).
Paper 4: Miocene sharks from Lesser Antilles
It wouldn’t be until 2008 that Gordon’s next paper (at least the next one recorded on Shark References) would be published, and it marked a return to the Caribbean. Namely, it is a paper led by Roger Portell of the Florida Museum that investigated a previously undocumented Miocene assemblage from Carriacou in the Grenadines – specifically in the Kendeace and Grand Bay formations (Portell et al. 2008).
Although three field trips were conducted, only five taxa were ultimately recovered: (1) the sand tiger shark (Carcharias taurus); (2) the shortfin mako shark (Isurus oxyrinchus); (3) the extinct megalodon (Otodus megalodon); (4) the dusky shark (Carcharhinus obscurus); and the extinct “wolverine shark” (Hemipristis serra) (Figure 8, 9). Curiously, the dusky shark was by far the most abundant of these fossils – having three times as many samples as all the other taxa combined (Portell et al. 2008). Today, the dusky shark lives in warm continental and insular shelf waters (Ebert et al. 2021) and so it would appear these habitats were important components of the Caribbean Miocene. Moreover, the presence of all the other taxa shows that this specific environment’s sharks harboured multiple ecological roles (Dedman et al. 2024) ranging from mesopredatory nutrient transporters (i.e., Williams et al. 2018) to high-level predators (i.e., Kent 1994) to super-predators like megalodon (Cooper et al. 2022; Kast et al. 2022; McCormack et al. 2022; 2025). Indeed, while megalodon was represented by just one tooth (Figure 8), it further suggests from Gordon’s earlier work that the Caribbean may well have been an important part of megalodon’s distribution, and perhaps of the wider Miocene shark community.
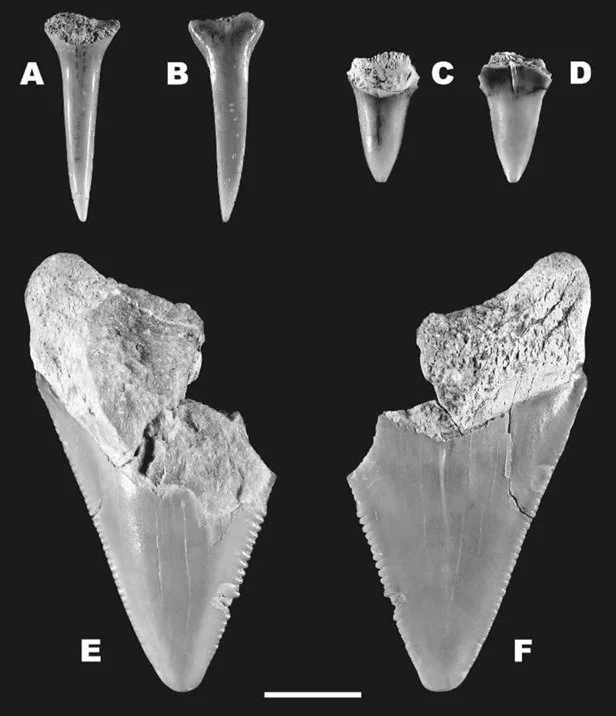
Figure 8. The teeth of (A-B) Carcharias taurus; (C-D) Isurus oxyrinchus; and (E-F) Otodus megalodon collected by the team in Gordon’s 2008 publication. Sourced from Figure 2 of Portell et al. (2008).
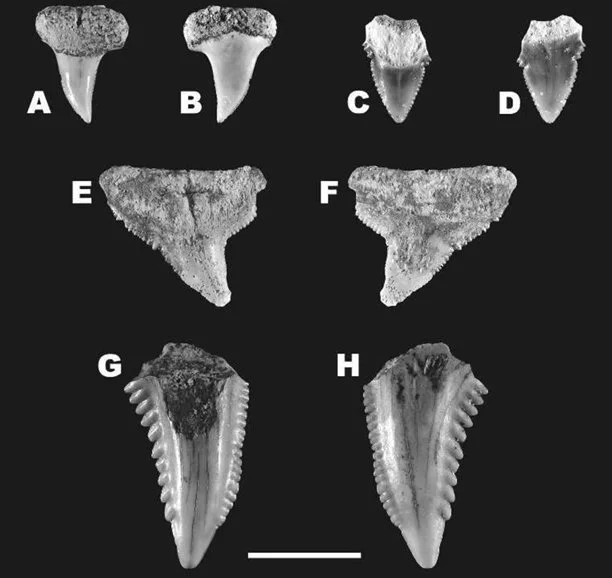
Figure 9. The teeth of (A-F) Carcharhinus obscurus; and (G-H) Hemipristis serra collected by the team of Gordon’s 2008 publication. Sourced from Figure 3 of Portell et al. (2008).
Paper 5: A truly exceptional fossil from an ancient white shark
Now this paper (Ehret et al. 2009) represents a key part of the PhD thesis by Dr Dana Ehret during his time working under Bruce MacFadden in Gainesville. It is also a paper that highlights probably the most famous fossil specimen from Gordon’s entire collection – specimen UF 226255 – consisting of jaws containing a whole dentition of 222 teeth, plus 45 associated vertebrae belonging to the genus Carcharodon from the Early Pliocene Pisco formation in Peru (Figure 10; Ehret et al. 2009). This fossil is so special because such preservation is extraordinarily rare. Shark skeletons only preserve under exceptional circumstances due to being made of soft cartilage (Hubbell 1996; Ehret et al. 2009). It had been discovered by Gordon in 1988 and only now was being described and added to the scientific record.
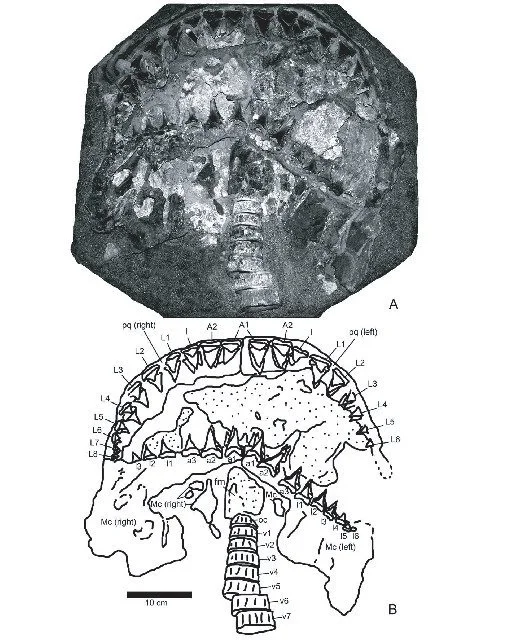
Figure 10. Photograph and schematic drawing of UF 226255; the holotype of Carcharodon hubbelli. See Figure 3 for my own photograph of the specimen. Sourced from Figure 4 of Ehret et al. (2009).
This particularly specimen, which would later be identified as an entirely new species of prehistoric white shark, Carcharodon hubbelli (named after Gordon himself; Ehret et al. 2012), was of great significance due to its role in resolving the debate around the evolution of the great white shark. At the time, researchers had debated for decades whether the modern great white shark had evolved either from ancient mako sharks or from megalodon. For well over a century, megalodon was deemed the great white’s direct ancestor, but this had been challenged in recent times. Most notably, geometric morphometrics of the teeth revealed distinct differences between the two species: namely, megalodon had thicker teeth, very different serrations and a dental band not seen in white shark teeth (Nyberg et al. 2006).
The Peruvian specimen, however, was an opportunity to assess an associated dentition and various aspects of anatomy not possible with isolated teeth alone. Specifically, Ehret et al. (2009) investigate not only the same tooth measurements as Hubbell (1996) but also calculate total length from these measurements and conduct x-rays and stable isotope analyses of the vertebrae preserved. Vertebral diameter measurements also allowed for growth calculations via growth bands and an alternative method for calculating total length.
The specimen was clearly Carcharodon due to having large symmetrical upper anterior teeth, a Carcharodon-style dentition and serrated teeth. However, the intermediate teeth were distally inclined as seen in ancient mako sharks. This led to Ehret et al. (2009) not declaring the specimen as a fossil of the living Carcharodon carcharias, paving the way for its eventual identification as the holotype of Carcharodon hubbelli (Ehret et al. 2012). Moreover, counting the vertebral growth bands revealed that the specimen was at least 20 years old, with oxygen-based stable isotope analyses supporting these bands being annually deposited. This ultimately indicated a slower growth than today’s great white shark, further evidence for this being a different species. Finally, the separate total length estimates put this specimen as belonging to a shark that was 4.8-5.1 m long (Ehret et al. 2009). Such an estimate would be supported by another methodology based on crown width, producing a size of 4.9 m in a later study (Perez et al. 2021).
The primitive mako shark traits combined with the derived white shark trait was further evidence of the white shark lineage emerging from ancient mako sharks (Ehret et al. 2009). Indeed, the species compared most to this specimen – named Isurus hastalis at the time – has since been revised as being a species of white shark: Carcharodon hastalis (Ehret et al. 2012), further supporting the closeness of these two taxa. As such, the old hypothesis that white sharks came from megalodon has been all but dismissed by the scientific community, with only a few rogue internet users standing by the old hypothesis in my experience, often ignoring the available evidence without explanation.
This incredible specimen of Carcharodon hubbelli was a key part of Gordon’s collection for decades and may well be the landmark fossil of his legacy; both for its remarkable preservation and for its eventual naming after him. It is thankfully deposited with a catalogue number with the Florida Museum – UF 226255 – making the specimen part of the museum’s public repository (Ehret et al. 2009; 2012). It was therefore available for researchers to study, something I hope will continue to be the case for decades to come. It was a true privilege to see this specimen up close with my own eyes during my visit to Gordon’s collection (Figure 10) and include it as data of my own work (Cooper & Pimiento 2024). I hope more work can continue on Carcharodon hubbelli, such as recent work finding teeth from New Zealand (Ehret et al. 2024).
Paper 6: The first ever nursery of megalodon
The next paper Gordon co-authored was published in 2010 as part of the masters work of another student of Bruce’s: Dr Catalina Pimiento, who would go to become my own PhD supervisor. And it is arguably one of the key studies that helped trigger a resurgence of academic work around the most iconic of fossil sharks: megalodon. Specifically, it identified the first ever documented nursery of the extinct giant shark (Pimiento et al. 2010).
Nurseries are a key part of the evolutionary success of sharks. They are typically shallow habitats filled primarily with baby sharks, but are defined by three key principles in living species: (1) sharks are more commonly encountered in these areas than others; (2) sharks either return frequently or remain in this area for prolonged periods; and (3) the area is repeatedly used over several years (Heupel et al. 2007). Now, it’s worth noting that these principles are virtually impossible to confirm in the inherently incomplete fossil record that operates on a geological time scale rather than an ecological one, and where isolated teeth are often all that remain of extinct sharks. As such, nurseries in the fossil record are identified primarily through evidence of a disproportionate presence of juveniles and geological evidence of a shallow, productive habitat. This evidence suggests that a certain fossil assemblage had both enough food for juvenile sharks to thrive and good shelter to be protected from larger sharks that might eat them (Pimiento et al. 2010; Herraiz et al. 2020; 2026). Prior to the study by Pimiento et al. (2010), only two fossil shark nurseries had reportedly been proposed (Purdy 1996; 1998).
This study investigated the Late Miocene Gatun formation in Panama, which had only had one descriptive study published prior (Gillette 1984). As part of her masters thesis (Pimiento 2010), Catalina had included palaeodepth and palaeoecology analyses that strongly indicated that the Gatun formation was a shallow, productive habitat –evidence that it could support a shark nursery. This would be further supported in a 2013 follow-up study that found that the area was filled with juveniles of various taxa (Pimiento et al. 2013). Although not common, megalodon teeth were found in Gatun and, curiously, most of them were small. As such, Catalina and her co-authors – Dana Ehret, Bruce MacFadden and of course Gordon Hubbell – set out to identify if Gatun was a fossil nursery of megalodon.
Collecting 28 megalodon teeth between 2007 and 2009 (Figure 11; even more small megalodon teeth would be collected as part of Pimiento et al. 2013), the team measured crown height and crown width. This allowed for calculations of body size using linear regressions from earlier work (Shimada 2003). The size of the teeth were compared with teeth from Maryland and Florida assemblages to determine if the sharks were significantly smaller than in other assemblages, and to complete dentitions from Gordon’s collection to identify approximate tooth position of each tooth (Pimiento et al. 2010). Using body size life stage cut-offs from Gottfried et al. (1996), the body size calculations from crown height indicated that the megalodons of Gatun were overwhelmingly juveniles (2-10.5 m in size; Figure 12). Although the life stage estimates are based entirely on extrapolations from the white shark, which have since been branded speculative with new calculations raising megalodon birth size (Perez et al. 2021; Shimada et al. 2025), the interpretation of disproportionately high representation of small megalodons holds due to how small the crown heights are (16-72 mm; Pimiento et al. 2010). As such, the conclusion was clear: Gatun appears to represent the first recorded nursery of megalodon (Pimiento et al. 2010).

Figure 11. The “baby shark” teeth of Otodus megalodon collected in the Gatun formation for Pimiento et al. (2010). On top of these 28 teeth, another 12 were collected as part of Pimiento et al. (2013) and another extra 12 are within the UF collections; with 48 of the 52 teeth I know of (i.e., 92.3%) coming from neonates or juveniles based on life stages from Gottfried et al. (1996; but see Perez et al. 2021; Shimada et al. 2025). When messaging the Pimiento group upon seeing these teeth myself at the Florida Museum, I jokingly referred to them as “Catalina’s baby [shark] teeth”. Sourced from Figure 2 of Pimiento et al. (2010).
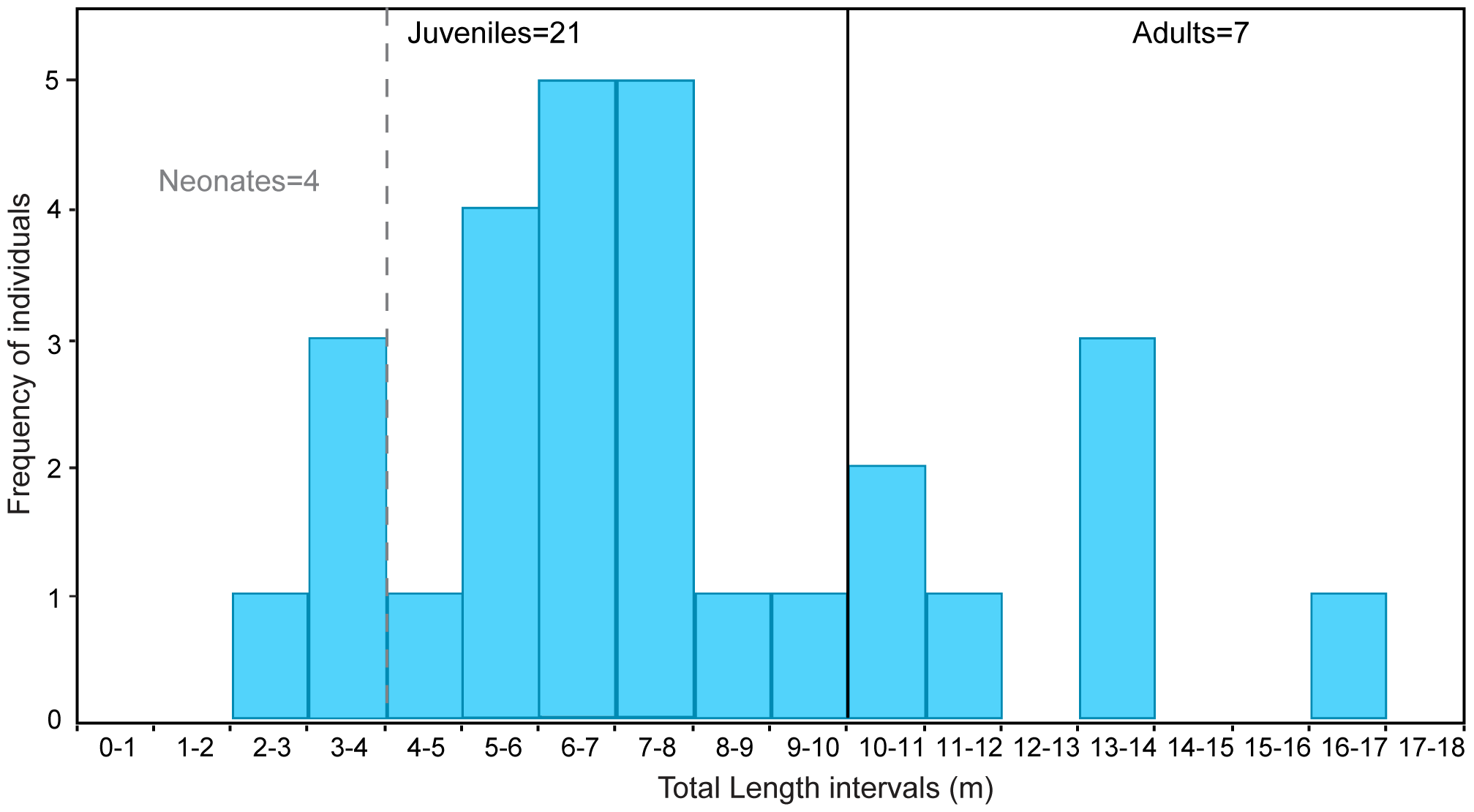
Figure 12. Histogram of the occurrence of megalodons at different life stages within the Gatun formation. Life stages based on Gottfried et al. (1996), but see Perez et al. (2021) for discussion on the validity of these cut-offs. The majority of the teeth clearly belong to juveniles based on these data; leading to the conclusion of Gatun as a megalodon nursery. Sourced from Figure 5 of Pimiento et al. (2010).
This study is significant for many reasons. Firstly, it kicked off a wave of new megalodon work driven primarily by Catalina’s PhD work (i.e., Pimiento & Clements 2014; Pimiento & Balk 2015; Pimiento et al. 2016). Since this period, there is at least one new megalodon paper every year across many different teams, significantly advancing our knowledge of the giant shark in too many areas and studies to list here. Secondly, it was the start of a recent series of studies that have identified and even debated fossil shark nurseries. In South America, fossil shark nurseries have been found for living species: the bronze whaler shark (Carcharhinus brachyurus) in the Late Miocene of Peru, and the great white shark (Carcharodon carcharias) from the Pliocene of Chile (Landini et al. 2017; Villafana et al. 2020). One of the earlier-studied fossil nursery sites (Purdy 1996) has now been identified as a nursery of megalodon’s ancestor Otodus angustidens (Miller et al. 2021). Fossil nurseries have even been identified for much older elasmobranch taxa from the Carboniferous and Triassic periods (Fischer et al. 2011; Sallan & Coates 2014).
And what about megalodon? Well, since the 2010 study, four additional sites have been proposed as megalodon nurseries across Panama, the United States and Spain (Herraiz et al. 2020). Although another study has argued that the body sizes across these sites may represent a case of climate-based Bergmann’s rule (where larger megs would have occupied colder waters and smaller megs would have lived in warmer waters; Shimada et al. 2022), a recent study has re-ignited this debate by arguing for the Spanish site’s status as a nursery (Herraiz et al. 2026). This debate will likely continue to unfold and as it does, I will be following it for a future blog on this website. But in the meantime, I can appreciate how this work co-authored by Gordon and masterminded by Catalina has been key in producing what may well be a current golden age of megalodon research.
Paper 7: Panamanian sharks from the Late Miocene and their functional diversity
This study is the third in a trio co-authored by Gordon and led by PhD students of Bruce MacFadden. In this case, Dr Victor Perez led a study that continues the theme from the last paper of Miocene sharks from Panama (Perez et al. 2017). In fact, it’s one of the most comprehensive studies of Neogene shark diversity in Panama, exploring one of the richest assemblages in the neotropics.
Investigating more than 1,400 chondrichthyan fossils from the Miocene Chucunaque Formation, the authors identified 23 shark taxa and 8 ray taxa across a wide range of habitats and feeding strategies. For example, the super-predator megalodon existed alongside smaller predators like mako and thresher sharks (Figure 13; Perez et al. 2017).
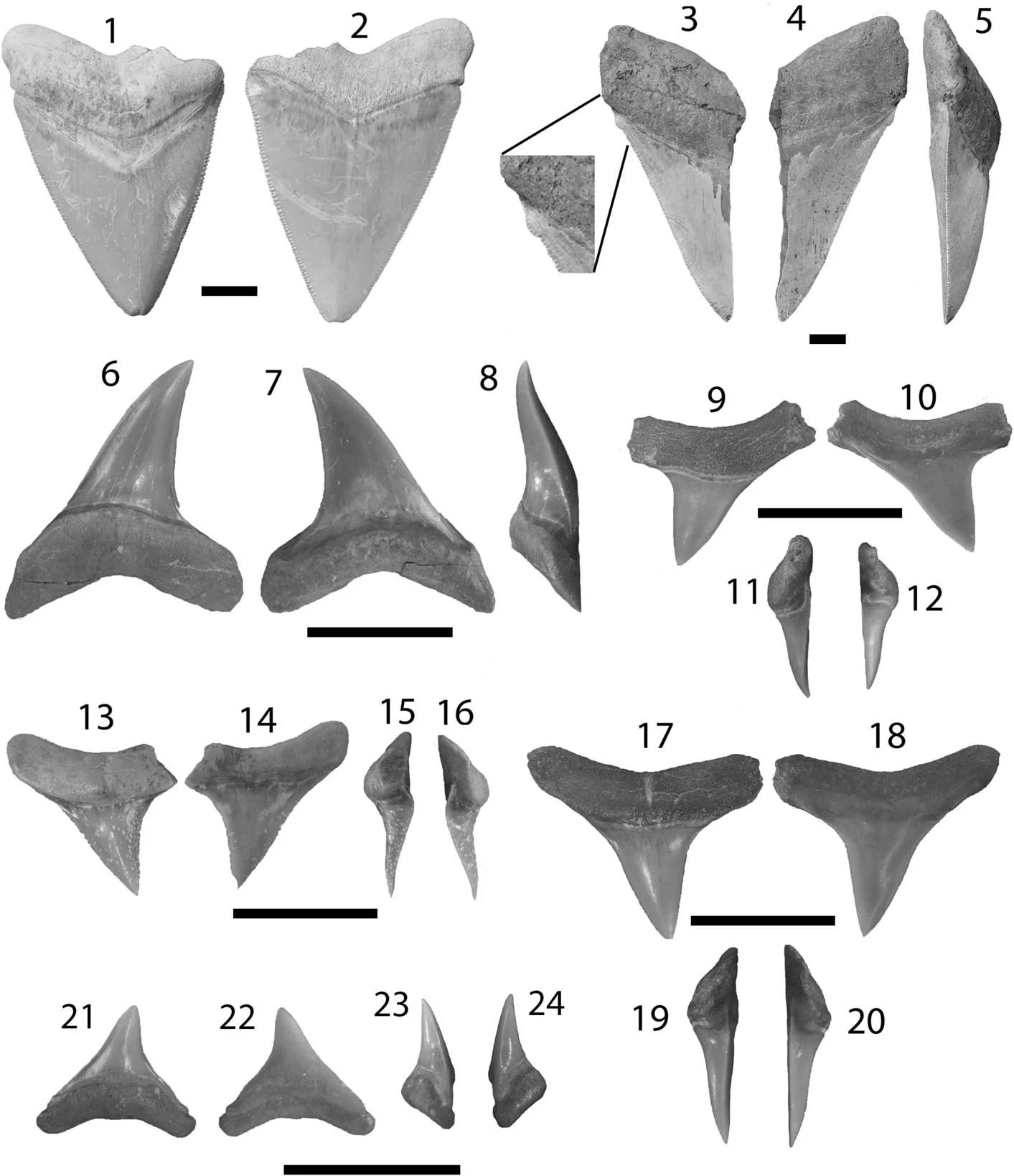
Figure 13. A subsample of the hundreds of teeth collected by the team from the Late Miocene of Panama. Sourced from Figure 4 of Perez et al. (2017).
This diversity of so many taxa indicates that, between 10 and 9.5 million years ago, the Chucunaque Formation was likely a highly productive marine ecosystem. Indeed, it probably supported high functional diversity (i.e., a large range of ecological roles; Mouillot et al. 2013; Cooper & Pimiento 2024), as shown by the range of different dentition types and ecomorphotypes (Figure 14). This ultimately indicates that, 10 million years ago, a strong community of sharks and rays occupied the waters of what would become Panama (Perez et al. 2017).
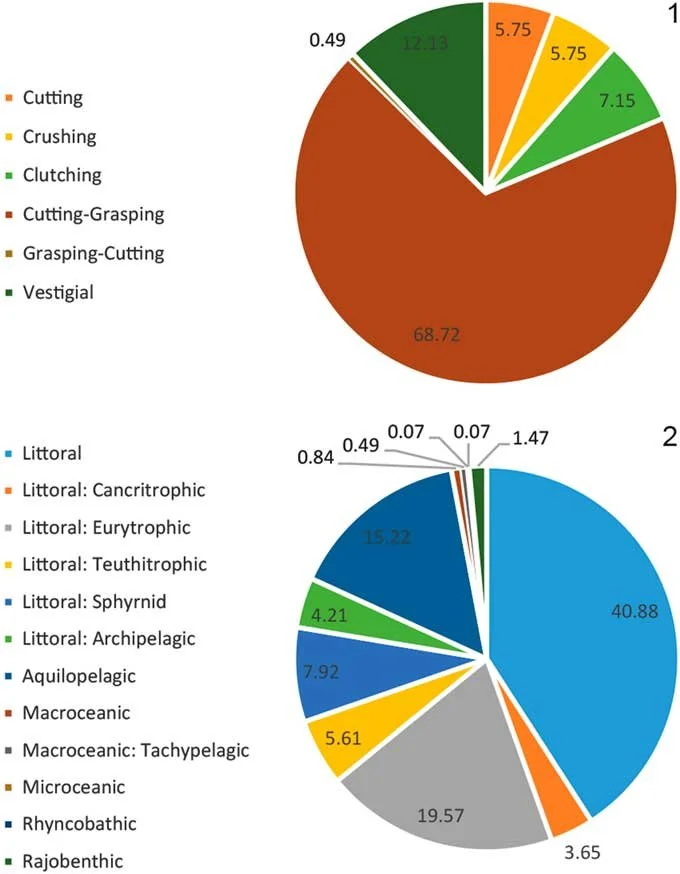
Figure 14. Proportional divisions of all the taxa collected by Perez et al. (2017) into different (1) dentition types, based on Kent (1994); and (2) ecomorphotypes, based on Compagno (1990). The range of both traits indicates a high functional diversity. Sourced from Figure 13 of Perez et al. (2017).
Notably, the Chucunaque Formation was deposited before the closure of the Isthmus of Panama, thought to have occurred around 3-3.5 million years ago (Bacon et al. 2015). This would have cut off the marine connections between the Caribbean and the Pacific Ocean. As such, like Gordon’s earlier works on the Caribbean (Iturralde-Vinent et al. 1996; Portell et al. 2008), this paper highlights that the Caribbean and Panamanian marine realms in the Miocene likely connected huge water basins, allowing species associated with all sorts of habitats to come through. As such, the paper arguably sets a high-quality baseline for evaluating faunal changes following the emergence of the Isthmus of Panama. By understanding how complex and diverse marine assemblages were before this significant geological event, we have a good starting point for evaluating the ecological and evolutionary shifts that likely came with it.
Paper 8: A comment on a mystery Miocene shark extinction event
Finally, we have a study co-authored by Gordon that made it to Science. Though this one is slightly different from the others in that it is a comment on another piece of work (Naylor et al. 2021). First, let’s just introduce a bit of background on a fascinating study that came out in 2021 and a subsequent debate around it.
Published in June of 2021, a paper in Science (Sibert & Rubin 2021a) reported that, 19 million years ago in the Early Miocene, an extinction event of unknown causes had suddenly occurred and hit pelagic sharks hard. Instead of teeth, Sibert and Rubin (2021a) collected dermal denticles (the microscopic tooth-like scales on shark skin) from deep-sea sediment cores and classified them as different morphotypes to reconstruct changes in shark diversity over time. Their analyses revealed that, 19 million years ago, there was a more than 70% decline in denticle morphotype diversity; with the abundance of those denticles also dropping by 90%. Importantly, both metrics did not recover following this event, leading to the authors interpreting this as a major, previously unknown, extinction event that remains unexplained (Sibert & Rubin 2021a).
Where Gordon comes in is that he was part of a comment that challenged this interpretation (Naylor et al. 2021). This comment made two particular critiques. Firstly, the authors questioned the relationship between denticle diversity and overall shark diversity. This is because modern sharks often possess several different denticle types across different regions of the body (Figure 15), and similar denticle types may occur in unrelated species. Secondly, Naylor et al. (2021) argued that changes in ocean circulation and habitat use can alter the denticle assemblage in deep-sea sediments without a dramatic extinction event. As such, the paper is ultimately arguing that denticle morphology across and within species must be better understood before making big assemblage-level conclusions; and that the observed declines may reflect ecological or geological changes rather than an extinction event (Naylor et al. 2021). It should be noted, however, that Sibert and Rubin responded to these criticisms in a follow-up work (Sibert & Rubin 2021b) in which, while acknowledging that denticles are not a perfect proxy for species diversity (let’s face it though; what is?), they ran bootstrap simulations that revealed an extinction of 90-93% of species in their huge dataset would have been needed to reproduce the estimated declines they saw in their original work.

Figure 15. Dermal denticle diversity across the entire body of one specimen of daggernose shark (Isogomphodon oxyrhynchus). All microscopic pictures of the denticles are scaled identically. Sourced from Figure 2A of Naylor et al. (2021).
Whichever side of the debate you fall under, both studies highlight the potential and limitations in the use of dermal denticles to reconstruct ancient shark communities. In the interest of a balanced conclusion, I think that something happened 19 million years ago that led to the observed declines, but the exact taxonomic magnitude of this event and thus how damaging it was cannot yet be precisely determined. I would love to see a follow-up to this work and for more research to dig into this mysterious event from 19 million years ago, whatever it was that happened.
Have any other papers used Gordon’s material?
As well as papers Gordon contributed to as a co-author, several other studies have used his material. In fact, it’s probably too many to list here so I’ll cover a few of my favourites.
One series of papers that aids our understanding of calculating body size of sharks from teeth is those by Professor Kenshu Shimada that constructed the linear relationships between crown height and body size in several different lamniform shark species. His work on the shortfin mako (Isurus oxyrinchus), great white shark (Carcharodon carcharias), sand tiger shark (Carcharias taurus), common thresher (Alopias vulpinus) and porbeagle shark (Lamna nasus) all utilised jaw specimens from Gordon’s collection – using 9, 10, 2, 3 and 12 jaw specimens respectively (Shimada 2002; 2003; 2004; 2006; Chavez et al. 2012). As such, Gordon’s collection has evidently been fundamental in establishing how we use teeth as ecological proxies.
Another key recent study that heavily used Gordon’s collection was another study by Dr Victor Perez, I believe initially conceived during his PhD with Bruce MacFadden. Specifically, this study (Perez et al. 2021) used multiple associated dentitions of megalodon, its ancestor Otodus chubutensis and multiple Carcharodon species, to come up with a new method of calculating body size. Crown height-based methods from the great white shark (Shimada 2003) had been shown to be effective when using upper anterior teeth in particular to conservatively calculate the size of megalodon (Shimada 2019), but when applied to associated dentitions across multiple tooth positions, the resulting total length estimates varied wildly (Grant et al. 2016). As such, Victor and his co-authors Ronny Leder and Teddy Badaut set out to use crown width instead. In short, the summed crown width (i.e., the combined crown width of all teeth in a row of an associated dentition; Figure 16) would be proportional to jaw width, which could then be proportional to total length (Perez et al. 2021). This method proved effective across multiple tooth positions and, when applied to the biggest megalodon tooth in Gordon’s collection, produced a maximum size estimate of 20 m (Perez et al. 2021; but see Shimada et al. 2025 for subsequent developments in megalodon body size). I remember speaking to Victor about how this study had come to be when I visited the Calvert Marine Museum in 2022 and he told me that Gordon had been invited to be a collaborator on the paper, but had been happy simply providing the material. There’s something I have to respect about providing the material and not necessarily needing any “credit” for the research that comes from it.
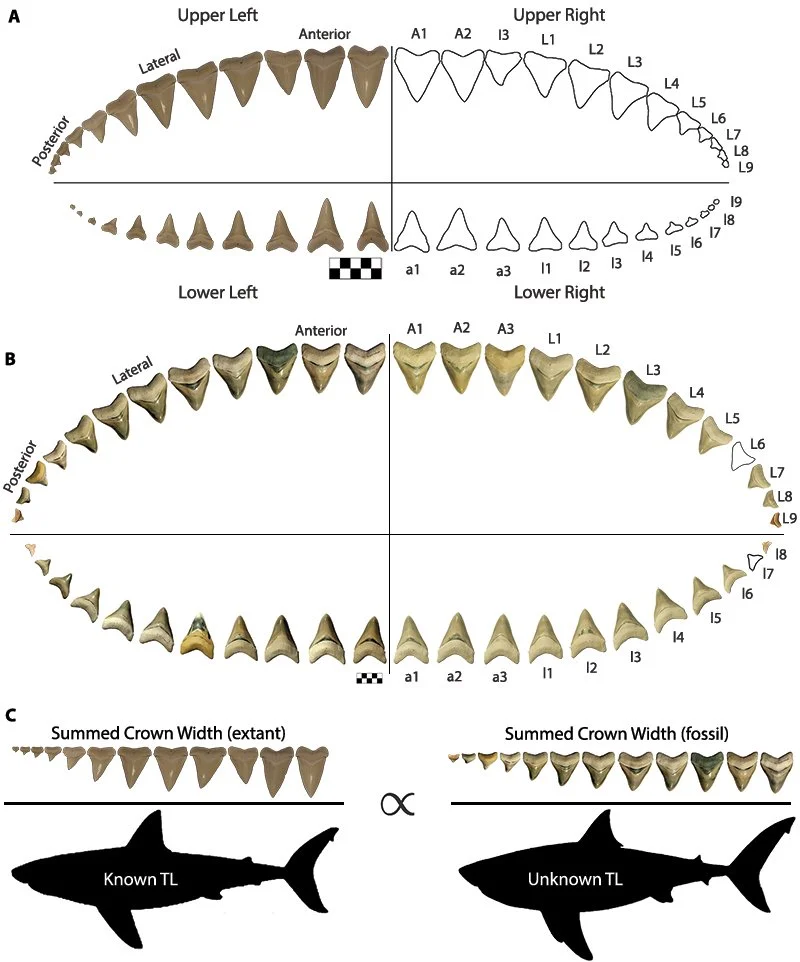
Figure 16. The basic concept of the methodology behind Perez et al. (2021). (A) A right-side dentition of the white shark from Hubbell (1996); (B) one of the most complete associated dentitions of megalodon in the entire fossil record, taken from Gordon’s collection; (C) the mathematical basis of the methodology applying summed crown widths from living sharks of known total length to extinct sharks. Sourced from Figure 1 of Perez et al. (2021).
Which brings us back to my own papers that, in part, used data from Gordon’s collection. To keep it short, the extant tooth and jaw specimens of various species in his collection were used as part of a “validation” test; investigating if dental characters could be used as proxies for functional traits in sharks (Cooper et al. 2023). A literature review had already identified over 60 shark taxa where this had been attempted across more than 50 studies. Museum specimens of these taxa, including several from Gordon’s collection, were then used to test for which dental characters drove the most variation in teeth and which best explained certain trait modalities (Figure 17). A fairly straightforward, perhaps even simplistic, concept for sure – but one that would have remained untested without Gordon’s collection.
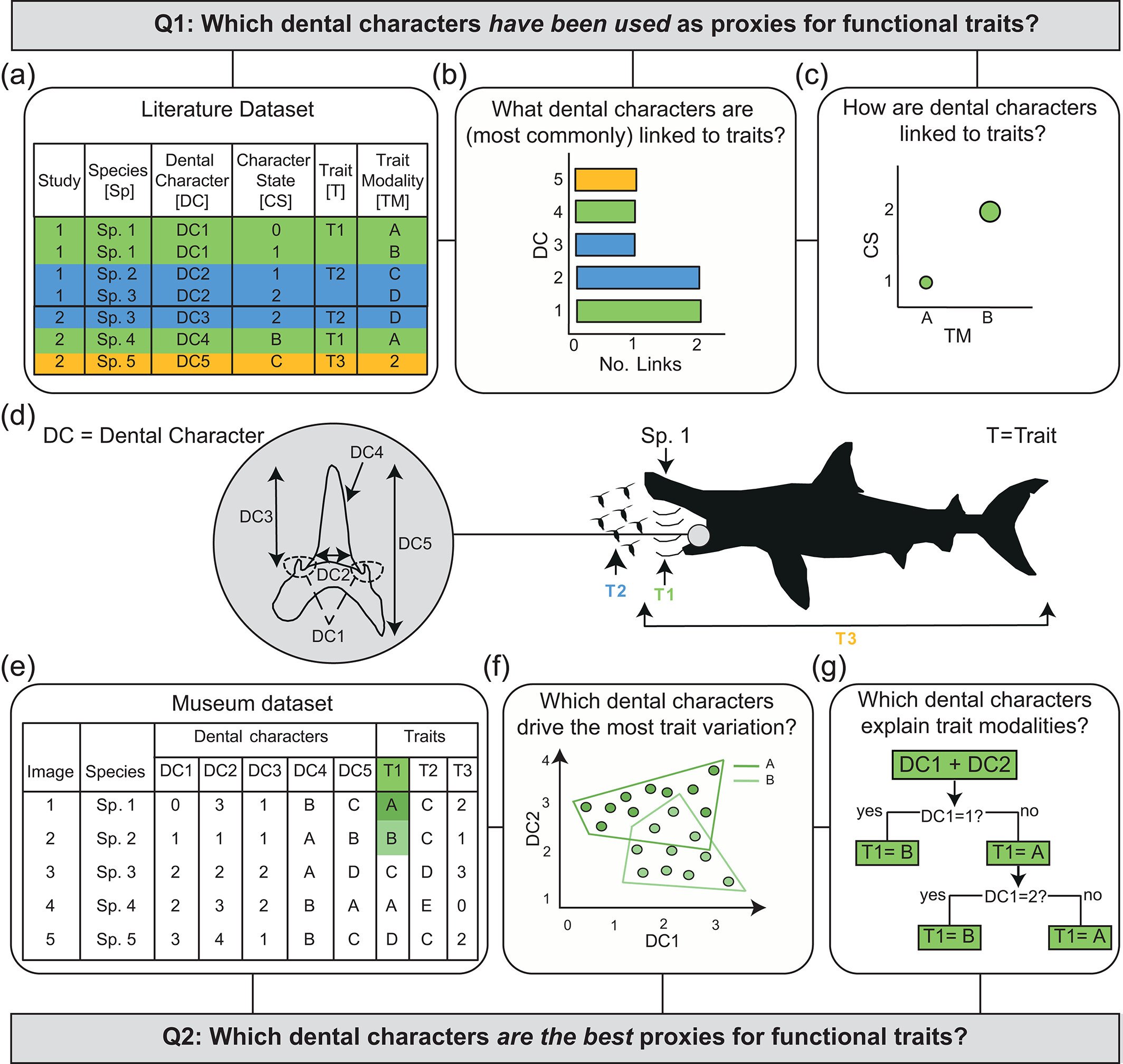
Figure 17. A schematic representation of the methodology behind the first chapter of my PhD. In (A-C), a literature review is assess where previous works have linked dental characters to functional traits (illustrated in D). In (E-G), the validation tests based on teeth from museum collections such as Gordon’s collection are illustrated. Sourced from Figure 1 of Cooper et al. (2023).
Following up from this first chapter (Cooper et al. 2023), I then collected the appropriate dental character measurements from over 9000 shark teeth spanning the entire Cenozoic era, including many from Gordon’s collection, to test how shark functional diversity changed during that era (Cooper & Pimiento 2024). Several species present in Gordon’s collection were not present in any of the other museums I visited, ensuring their inclusion for a more complete analysis. Gordon was acknowledged for providing access to his collection in both of my papers and I cannot emphasise enough how important that access was for my work to be as well-sampled as possible.
I remain extremely grateful for that, and for the wonderful hospitality Gordon provided during my visit.
Concluding reflections
When we look back on Gordon’s legacy, we of course look at the remarkable collection of shark specimens and fossils he acquired over his many decades. That should of course not be taken for granted, nor should the fact that this collection was made so readily available to researchers. Having now dived into the academic works he contributed to, however, I think it also gives a perspective as to the major advances in shark science he helped drive forward. A few recurring themes pop up across even in just this small sample of eight papers.
One theme that stands out is how shark scientists must understand variation within the fossils of individual shark species before making any evolutionary or ecological conclusions. This is most clearly seen in Hubbell (1996) where white shark teeth were seen to vary between individual positions and across ontogeny but also resurfaced in his final paper (Naylor et al. 2021) looking at variation of dermal denticles within the same shark species (Figure 15). In short, we can only make so many conclusions from small samples. We must be cautious in our interpretations (e.g., naming new species from a small sample of teeth) and rigorous in understanding if tooth measurements that might inform ecology in shark fossils could in any way be influenced by jaw position or ontogeny. Indeed, in the first paper of my PhD (Cooper et al. 2023), tooth position was considered as part of the analyses for this very reason.
A second theme I spotted across Gordon’s publications was the use of shark fossils as proxies for ancient ecosystems, particularly around the Caribbean and Panama. The Cuban shark fossil record (Iturralde-Vinent et al. 1996) was used to not only establish a more complete diversity of sharks from the Neogene in this region but also to infer the existence of a seaway through western Cuba, showing how fossil sharks reveal ecosystem structure. This theme would be revisited in Portell et al. (2008) and also explored in how rich Miocene shark assemblages from Panama reveal ecosystem structure prior to the formation of the Isthmus of Panama in Perez et al. (2017). And of course, we can’t forget about the proposed megalodon nursery in Pimiento et al. (2010) showcasing how the giant shark may have used the Panamanian waters in the very beginning of its life.
Thirdly, Gordon’s work often used information from modern sharks and connected it with the fossil record. His work with Kenshu Shimada in Shimada & Hubbell (2001) connected abnormal teeth in the extinct Cretoxyrhina with those in living Alopias to interpret similar conclusions separated by millions of years. Meanwhile, the modern concept of shark nurseries was applied directly to megalodon in Pimiento et al. (2010). In short, these studies showcased how living sharks can be used as analogues for understanding extinct species.
A final important theme spanning at least two of Gordon’s papers was the evolution of the great white shark. The study of teeth in Hubbell (1996) was ultimately all about understanding how teeth can be used to study the evolution of this most iconic of living sharks. And of course, the exceptional specimen of Carcharodon hubbelli detailed in Ehret et al. (2009) was pivotal data for supporting the hypothesis that white sharks did not evolve from megalodon as originally thought, but instead from ancient makos. It was so crucial because its remarkable preservation provided so much more information than isolated teeth could ever do.
Taken all together, Gordon’s academic legacy is not just in acquiring his wonderful collection, but also in demonstrating how carefully documented specimens can recreate shark evolution, reconstruct ancient ecosystems and connect the biology of living sharks to the fossil record of their extinct counterparts. But perhaps Gordon's greatest contribution was not simply the specimens he collected, but the way he made them available to others. I hope that Gordon’s collection will not be sold to the highest bidder, but instead deposited within the Florida Museum’s repository, or in another public repository, following his passing. That way, his remarkable specimens will not be lost to science; a real challenge facing our field when it comes to private collections and fossil commercialisation (Shimada et al. 2014).
Whatever happens next, the impact that Gordon has made on shark science cannot be understated. I will remember my discovery of his collection as a child and my interactions with him fondly, particularly meeting him in person on that hot humid day in Gainesville in 2022. But I will also reflect on just how much we have learned thanks to his passion for the field we all love and the collection he built from that. He will undoubtedly be dearly missed in our little community of fossil shark researchers and enthusiasts alike (Figure 18).
May his legacy continue.

Figure 18. A graphic that has made the rounds amongst groups of shark enthusiasts on social media in the days since Gordon’s passing.
References
Bacon CD, Silvestro D, Jaramillo C, Smith BT, Chakrabarty P & Antonelli A, 2015. Biological evidence supports an early and complex emergence of the Isthmus of Panama. Proceedings of the National Academy of Sciences, 112, 6110-6115.
Bemis WE, Moyer JK & Riccio ML, 2015. Homology of lateral cusplets in the teeth of lamnid sharks (Lamniformes: Lamnidae). Copeia, 103, 961-972.
Chavez S, Zufan S, Kim SH & Shimada, K., 2012. Tooth sizes as a proxy for estimating body lengths in the porbeagle shark, Lamna nasus. Journal of Fossil Research, 45, 1-5.
Compagno LJV, 1990. Alternative life-history styles of cartilaginous fishes in time and space. Environmental Biology of Fishes, 28, 33-75.
Cooper JA, Griffin JN, Kindlimann R & Pimiento C, 2023. Are shark teeth proxies for functional traits? A framework to infer ecology from the fossil record. Journal of Fish Biology, 103, 798-814.
Cooper JA, Hutchinson JR, Bernvi DC, Cliff G, Wilson RP, Dicken ML, Menzel J, Wroe S, Pirlo J & Pimiento C, 2022. The extinct shark Otodus megalodon was a transoceanic superpredator: Inferences from 3D modeling. Science Advances, 8, eabm9424.
Cooper JA & Pimiento C, 2024. The rise and fall of shark functional diversity over the last 66 million years. Global Ecology and Biogeography, 33, e13881.
Dedman S, Moxley JH, Papastamatiou YP, Braccini M, Caselle JE, Chapman DD, Cinner JE, Dillon EM, Dulvy NK, Dunn RE, Espinoza M, Harborne AR, Harvey ES, Heupel MR, Huvenners C, Graham NAJ, Ketchum JT, Klinard NV, Kock AA, Lowe CG, MacNeil MA, Madin EMP, McCauley DJ, Meekan MG, Meier AC, Simpfendorfer CA, Tinker MT, Winton M, Wirsing AJ & Heithaus MR, 2024. Ecological roles and importance of sharks in the Anthropocene Ocean. Science, 385, adl2362.
Eastman CR, 1895. Beiträge zur Kenntniss der Gattung Oxyrhina mit besonderer Berücksichtigung von Oxyrhina mantelli Agassiz. Palaeontographica, 41, 149–191.
Ebert DA, Dando M & Fowler S, 2021. Sharks of the world: a complete guide. Princeton University Press.
Ehret DJ, Hubbell G & MacFadden BJ, 2009. Exceptional preservation of the white shark Carcharodon (Lamniformes, Lamnidae) from the early Pliocene of Peru. Journal of Vertebrate Paleontology, 29, 1-13.
Ehret DJ, MacFadden BJ, Jones DS, Devries TJ, Foster DA & Salas-Gismondi, R, 2012. Origin of the white shark Carcharodon (Lamniformes: Lamnidae) based on recalibration of the Upper Neogene Pisco Formation of Peru. Palaeontology, 55, 1139-1153.
Ehret DJ, Tennyson AJ, Richards MD & Boessenecker RW, 2024. First records of two mackerel shark species (Carcharodon planus comb. nov. and Carcharodon hubbelli; Lamnidae) from New Zealand. Journal of the Royal Society of New Zealand, 54, 609-619.
Fischer J, Voigt S, Schneider JW, Buchwitz M & Voigt S, 2011. A selachian freshwater fauna from the Triassic of Kyrgyzstan and its implication for Mesozoic shark nurseries. Journal of Vertebrate Paleontology, 31, 937-953.
French GCA, Stürup M, Rizzuto S, Van Wyk JH, Edwards D, Dolan RW, Wintner SP, Towner AV & Hughes WOH, 2017. The tooth, the whole tooth and nothing but the tooth: tooth shape and ontogenetic shift dynamics in the white shark Carcharodon carcharias. Journal of Fish Biology, 91, 1032-1047.
Gillette DD, 1984. A marine ichthyofauna from the Miocene of Panama, and the Tertiary Caribbean faunal province. Journal of Vertebrate Paleontology, 4, 172-186.
Gottfried MD, Compagno LJV & Bowman SC, 1996. Size and skeletal anatomy of the giant megatooth shark Carcharodon megalodon. In AP Klimley & DG Ainley, (Eds.), Great white sharks: the biology of Carcharodon carcharias. Academic Press, pp. 55-66.
Grant CA, MacFadden BJ, Antonenko P & Perez VJ, 2016. 3-D fossils for k–12 education: A case example using the giant extinct shark Carcharocles megalodon. The Paleontological Society Papers, 22, 197-209.
Herraiz JL, Ferrón HG, Botella H, Reolid M & Martínez-Pérez C, 2026. The Iberian fossil record of †Otodus megalodon rejects Mediterranean dwarfism and supports nursery use. Biology Letters, 22, 20250640.
Herraiz JL, Ribé J, Botella H, Martínez-Pérez C & Ferrón HG, 2020. Use of nursery areas by the extinct megatooth shark Otodus megalodon (Chondrichthyes: Lamniformes). Biology Letters, 16, 20200746.
Heupel MR, Carlson JK & Simpfendorfer CA, 2007. Shark nursery areas: concepts, definition, characterization and assumptions. Marine Ecology Progress Series, 337, 287-297.
Hubbell G, 1996. Using tooth structure to determine the evolutionary history of the white shark. In AP Klimley & DG Ainley, (Eds.), Great white sharks: the biology of Carcharodon carcharias. Academic Press, pp. 9-18.
Iturralde-Vinent M, Hubbell G & Rojas R, 1996. Catalog of Cuban fossil Elasmobranchii (Paleocene—Pliocene) and paleooceanographic implications of their lower—middle Miocene occurrence. Boletín de la Sociedad Jamaicana de Geología, 31, 7-21.
Kast ER, Griffiths ML, Kim SL, Rao ZC, Shimada K, Becker MA, Maisch HM, Eagle RA, Clarke CA, Neumann AN, Karnes ME, Lüdecke T, Leichliter JN, Martínez-García A, Akhtar AA, Wang XT, Haug GH & Sigman DM, 2022. Cenozoic megatooth sharks occupied extremely high trophic positions. Science Advances, 8, eabl6529.
Kent BW, 1994. Fossil sharks of the Chesapeake Bay region. Egan Rees & Boyer, Inc.
Landini W, Collareta A, Pesci F, Di Celma C, Urbina M & Bianucci G, 2017. A secondary nursery area for the copper shark Carcharhinus brachyurus from the late Miocene of Peru. Journal of South American Earth Sciences, 78, 164-174.
McCormack J, Feichtinger I, Fuller BT, Jaouen K, Griffiths ML, Bourgon N, Maisch IV H, Becker MA, Pollerspöck J, Hampe O, Rössner GE, Assemat A, Müller W & Shimada K, 2025. Miocene marine vertebrate trophic ecology reveals megatooth sharks as opportunistic supercarnivores. Earth and Planetary Science Letters, 664, 119392.
McCormack J, Griffiths ML, Kim SL, Shimada K, Karnes M, Maisch H, Pederzani S, Bourgon N, Jaouen K, Becker MA, Jöns N, Sisma-Ventura G, Straube N, Pollerspöck J, Hublin JJ, Eagle RA & Tütken T, 2022. Trophic position of Otodus megalodon and great white sharks through time revealed by zinc isotopes. Nature Communications, 13, 2980.
Miller AE, Gibson ML & Boessenecker RW, 2021. A megatoothed shark (Carcharocles angustidens) nursery in the Oligocene Charleston Embayment, South Carolina, USA. Palaeontologia Electronica, 24.2, a19.
Mouillot D, Graham NA, Villéger S, Mason NW & Bellwood DR, 2013. A functional approach reveals community responses to disturbances. Trends in Ecology & Evolution, 28, 167-177.
Naylor GJ, de Lima A, Castro JI, Hubbell G & de Pinna MC, 2021. Comment on "An early Miocene extinction in pelagic sharks". Science, 374, eabj8723.
Nyberg KG, Ciampaglio CN & Wray GA, 2006. Tracing the ancestry of the great white shark, Carcharodon carcharias, using morphometric analyses of fossil teeth. Journal of Vertebrate Paleontology, 26, 806-814.
Perez VJ, Leder RM & Badaut T, 2021. Body length estimation of Neogene macrophagous lamniform sharks (Carcharodon and Otodus) derived from associated fossil dentitions. Palaeontologia Electronica, 24, a09.
Perez VJ, Pimiento C, Hendy A, González-Barba G, Hubbell G & MacFadden BJ, 2017. Late Miocene chondrichthyans from Lago Bayano, Panama: functional diversity, environment and biogeography. Journal of Paleontology, 91, 512-547.
Pimiento C, 2010. Systematics, paleobiology, and paleoecology of late Miocene sharks (Elasmobranchii, Selachii) from Panama: integration of research and education. Gainesville: University of Florida. 131 p.
Pimiento C & Balk MA, 2015. Body-size trends of the extinct giant shark Carcharocles megalodon: a deep-time perspective on marine apex predators. Paleobiology, 41, 479-490.
Pimiento C & Clements CF, 2014. When did Carcharocles megalodon become extinct? A new analysis of the fossil record. PLoS One, 9, e111086.
Pimiento C, Ehret DJ, MacFadden BJ & Hubbell G, 2010. Ancient nursery area for the extinct giant shark Megalodon from the Miocene of Panama. PLoS One, 5, e10552.
Pimiento C, González-Barba G, Ehret DJ, Hendy AJ, MacFadden BJ & Jaramillo C, 2013. Sharks and rays (Chondrichthyes, Elasmobranchii) from the late Miocene Gatun formation of Panama. Journal of Paleontology, 87, 755-774.
Pimiento C, MacFadden BJ, Clements CF, Varela S, Jaramillo C, Velez‐Juarbe J & Silliman BR, 2016. Geographical distribution patterns of Carcharocles megalodon over time reveal clues about extinction mechanisms. Journal of Biogeography, 43, 1645-1655.
Portell RW, Hubbell G, Donovan SK, Green JL, Harper DA & Pickerill R, 2008. Miocene sharks in the Kendeace and Grand Bay formations of Carriacou, The Grenadines, Lesser Antilles. Caribbean Journal of Science, 44, 279-286.
Purdy RW, 1996. Paleoecology of fossil white sharks. In AP Klimley & DG Ainley, (Eds.), Great white sharks: the biology of Carcharodon carcharias. Academic Press, pp. 67-78.
Purdy RW, 1998. Chondrichthyan fishes from the Paleocene of South Carolina. Transactions of the American Philosophical Society, 88, 122-146.
Sallan LC & Coates MI, 2014. The long-rostrumed elasmobranch Bandringa Zangerl, 1969, and taphonomy within a Carboniferous shark nursery. Journal of Vertebrate Paleontology, 34, 22-33.
Shimada K, 1997a. Dentition of the Late Cretaceous lamniform shark, Cretoxyrhina mantelli from the Niobrara Chalk of Kansas. Journal of Vertebrate Paleontology, 17, 269-279.
Shimada K, 1997b. Skeletal anatomy of the Late Cretaceous lamniform shark, Cretoxyrhina mantelli from the Niobrara Chalk in Kansas. Journal of Vertebrate Paleontology, 17, 642-652.
Shimada K, 1999. Contribution of dental characters to elucidate the phylogeny of lamniform sharks. Journal of Vertebrate Paleontology, 19, 75A-76A.
Shimada K, 2002. The relationship between the tooth size and total body length in the shortfin mako, Isurus oxyrinchus (Lamniformes: Lamnidae). Journal of Fossil Research, 35, 6-9.
Shimada K, 2003. The relationship between the tooth size and total body length in the white shark, Carcharodon carcharias (Lamniformes: Lamnidae). Journal of Fossil Research, 35, 28-33.
Shimada K., 2004. The relationship between the tooth size and total body length in the sandtiger shark, Carcharias taurus (Lamniformes: Odontaspididae). Journal of Fossil Research, 37, 76-81.
Shimada K, 2006. The relationship between the tooth size and total body length in the common thresher shark, Alopias vulpinus (Lamniformes: Alopiidae). Journal of Fossil Research, 39, 7-11.
Shimada K, 2019. The size of the megatooth shark, Otodus megalodon (Lamniformes: Otodontidae), revisited. Historical Biology, 33, 904-911.
Shimada K, Chandler RE, Lam OLT, Tanaka T & Ward DJ, 2017. A new elusive otodontid shark (Lamniformes: Otodontidae) from the lower Miocene, and comments on the taxonomy of otodontid genera, including the ‘megatoothed’ clade. Historical Biology, 29, 704-714.
Shimada K, Currie PJ, Scott E & Sumida SS, 2014. The greatest challenge to 21st century paleontology: When commercialization of fossils threatens the science. Palaeontologia Electronica, 17, 1E.
Shimada K & Hubbell G, 2001. Identity of small symmetrical teeth of the Late Cretaceous lamniform shark, Cretoxyrhina mantelli, from western Kansas, USA. Journal of Fossil Research, 34, 55-57.
Shimada K, Maisch IV HM, Perez VJ, Becker MA & Griffiths ML, 2022. Revisiting body size trends and nursery areas of the Neogene megatooth shark, Otodus megalodon (Lamniformes: Otodontidae), reveals Bergmann’s rule possibly enhanced its gigantism in cooler waters. Historical Biology, 35, 208-217.
Shimada K, Motani R, Wood JJ, Sternes PC, Tomita T, Bazzi M, Collareta A, Gayford JH, Türtscher J, Jambura PL, Kriwet J, Vullo R, Long DJ, Summers AP, Maisey JG, Underwood C, Ward DJ, Maisch HM, Perez VJ, Feichtinger I, Naylor GJP, Moyer JK, Higham TE, da Silva JPCB, Bornatowski H, González-Barba G, Griffiths ML, Becker MA & Siversson M, 2025. Reassessment of the possible size, form, weight, cruising speed, and growth parameters of the extinct megatooth shark, Otodus megalodon (Lamniformes: Otodontidae), and new evolutionary insights into its gigantism, life history strategies, ecology, and extinction. Palaeontologia Electronica, 28, a12.
Sibert EC & Rubin LD, 2021a. An early Miocene extinction in pelagic sharks. Science, 372, 1105-1107.
Sibert EC & Rubin LD, 2021b. Response to comment on "An early Miocene extinction of pelagic sharks". Science, 374, eabj9522.
Sternes PC, Jambura PL, Türtscher J, Kriwet J, Siversson M, Feichtinger I, Naylor GJ, Summers AP, Maisey JG, Tomita T, Moyer JK, Higham TE, da Silva JPCB, Bornatowski H, Long DJ, Perez VJ, Collareta A, Underwood C, Ward DJ, Vullo R, González-Barba G, Maisch HM, Griffiths ML, Becker MA, Wood JJ & Shimada K, 2024. White shark comparison reveals a slender body for the extinct megatooth shark, Otodus megalodon (Lamniformes: Otodontidae). Palaeontologia Electronica, 27, a7.
Villafaña JA, Hernandez S, Alvarado A, Shimada K, Pimiento C, Rivadeneira MM & Kriwet J, 2020. First evidence of a palaeo-nursery area of the great white shark. Scientific Reports, 10, 8502.
Williams JJ, Papastamatiou YP, Caselle JE, Bradley D & Jacoby DM, 2018. Mobile marine predators: an understudied source of nutrients to coral reefs in an unfished atoll. Proceedings of the Royal Society B: Biological Sciences, 285, 20172456.