Megalodon and its relatives were super-predators that ate whatever they wanted
The iconic megalodon shark is generally pictured as having been a specialised whale killer. Now, a new study finds that megalodon and its closest relative Otodus chubutensis weren’t picky eaters. Instead, they consumed not only marine mammals, but a wide range of prey across populations.
The traditional image of megalodon; and what we’ve learned about its trophic level
What’s the first thing you picture when you think of Otodus megalodon?
Perhaps it’s the gigantic size. After all it was the biggest shark that ever lived (Pimiento et al. 2024) – reaching maximum sizes of at least 18-20 m long (Pimiento & Balk 2015; Cooper et al. 2020; Perez et al. 2021) and maybe even as big as 24 m (Shimada et al. 2025; see also my blog on that paper). For others, however, they think of this enormous shark munching down on a whale. You might imagine the shark shaking its head to rip apart the stricken whale. Or, perhaps, the huge predator leaping out of the water with a whale in its mouth in similar vein to breaching predations seen in the living great white shark (Carcharodon carcharias) snatching a seal in South Africa (Klimley 1994; Martin et al. 2005).
While the breaching behaviour is speculation on my part, as no fossil evidence has ever been found to suggest such a predatory tactic for megalodon, the image of the massive animal feasting on whales is not unfounded (Figure 1). We have plenty of evidence that megalodon ate whales and other marine mammals – specifically in trace fossils where the shark’s bite marks are left behind on the bones. Bite marks have been found on marine mammal vertebrae, ribs, tail bones and even teeth (Godfrey & Altman 2005; Aguilera et al. 2008; Kallal et al. 2010; Collareta et al. 2017; Godfrey et al. 2018; 2021; Godfrey & Beatty 2022). This includes some fossils with megalodon teeth still stuck inside (Aguilera et al. 2008); and one fossil prey has even been identified to species-level: the small baleen whale Piscobalena nana (Collareta et al. 2017; Figure 1A). With one of the strongest bite forces in history, megalodon would’ve caused devastating damage to its prey (Wroe et al. 2008), and this is reflected in some of the most fascinating bite mark fossils… ones where the prey has miraculously survived the bite! Some whale bones have been found with evidence of healing (Godfrey & Altman 2005; Kallal et al. 2010; Godfrey & Beatty 2022; Figure 1B, C), clearly showing that megalodon was very much an active predator that would occasionally fail, just like all predators. So what were megalodon’s menu options if it failed to eat a tasty whale, and what precisely was its trophic level?

Figure 1. Palaeoartistic depictions of Otodus megalodon predations on whales and the “bite mark” trace fossils left behind. (A) a predation of the small baleen whale Piscobalaena nana; sourced from the graphical abstract of Collareta et al. (2017), with artwork by Alberto Gennari; (B) a breaching predation of a baleen whale; artwork by Clarence Schumaker; and (C) the damaged vertebra left behind showing evidence of healing; studied by Godfrey & Beatty (2022). Panels B and C are sourced from Bird (2022).
As it happens, several recent studies have suggested that there was more to megalodon’s diet than simply being a big whale killer. One particularly interesting fossil bite mark was on the tooth of a raptorial sperm whale (Godfrey et al. 2021), indicating that megalodon could eat other predators rather than just baleen whales. My own work making a 3D model of megalodon based on the most complete vertebral column further suggested a stomach volume that would allow the shark to feast on prey as large as 8 m, as big as today’s killer whale (Cooper et al. 2022), supporting such a notion. This implied that megalodon was on a higher trophic level than any marine predator alive today. It would therefore have been an “apex super-predator” as my paper described at the time, though another name for this would be “supercarnivore”.
While these results gave clear implications, a recent series of very clever papers has produced more direct, data-driven, evidence for megalodon’s diet. These papers all share a common theme: the use of stable isotopes. To put it simply (chemistry was never my strongest subject at school), stable isotopes of certain elements vary in ratio and composition in teeth depending on food source. Therefore, by extracting these isotopes and analysing them, scientists can infer both what an animal was eating and its trophic level. These isotopes incredibly preserve within fossils, meaning that this can be directly applied to the fossil record too (Casey & Post 2011). In the case of megalodon, a 2015 paper extracting calcium isotopes indicated that its trophic level was higher than that of the living great white shark (Martin et al. 2015). In 2022, another two studies were published that offered intriguing results – one using zinc isotopes found that megalodon’s trophic level was comparable to the white shark (McCormack et al. 2022), while the other used nitrogen isotopes and found that megalodon and its ancestors dating as far back as the Eocene had trophic levels higher than any living marine predator (Kast et al. 2022). Complicating matters further was dental microwear analyses recently finding no significant difference between megalodon teeth and those of the white shark in wear patterns, suggesting similar diets and therefore trophic overlap (Paredes-Aliaga & Herraiz 2024). If the white shark truly was at a similar trophic level to megalodon then it would add credence to the hypothesis that competition between the two may have played a role in megalodon’s eventual extinction (Pimiento et al. 2016; Boessenecker et al. 2019; McCormack et al. 2022; Shimada et al. 2025). But if that’s true, why are there also other studies suggesting megalodon was a supercarnivore at a higher trophic level than anything in our ocean today?
So, to summarise the key lingering question, was megalodon a supercarnivore or was it at a similar trophic level to the living white shark? This is where a fascinating new study (McCormack et al. 2025) comes in. Led by Dr Jeremy McCormack, who led one of the 2022 studies, and featuring an ensemble of expert fossil shark researchers, a re-application of the zinc isotope application may have cracked the case.
The new study
209 teeth from 19 different marine taxa were gathered – 150 new specimens and 59 already published – for zinc isotope extractions: with lower values generally indicating higher trophic levels (McCormack et al. 2025). Notably, the fossil specimens were all gathered from the Early Miocene of southern Germany in only a select few geological formations, meaning that the results would provide a snapshot of the marine food web in this area between 20 and 15 million years ago. In other words, the authors are capturing the marine food web of a whole area at a single point in deep time.
To obtain the zinc isotopes, enameloid was drilled out of the teeth, and the zinc was then purified with ion exchange chromatography and measured. Size estimates for megalodon and its relative Otodus chubutensis, as well as Carcharodon hastalis, the ancestor of the great white shark (Ehret et al. 2012), were all made using crown height based on tooth position and linear equations from white sharks (Shimada 2003). Finally, differences between zinc isotopes across individual species – and between the same species in different localities – were assessed with ANOVA and Tukey post-hoc tests. The latter tests were particularly interesting as they allow the authors to not only assess if different species occupy different trophic levels, but if the same individual species may vary in diet and trophic level between populations; something already known to occur in living sharks (Assemat et al. 2022).
The results produced by these analyses are fascinating. The zinc isotope ranges had a clear trophic stratification (Figure 2), essentially capturing multiple marine trophic levels of the German Early Miocene. Moreover, zinc isotope values of fossil specimens – be it sharks or toothed whales – overlapped with living analogues; giving a clear control suggesting that the fossil values were accurate.
Ultimately, there are three key results that stand out here to me. Firstly, zinc values are lowest in the Otodus species – specifically megalodon and Otodus chubutensis – and this clearly indicates that they could occupy the highest trophic levels. Simultaneously, however, their range of zinc isotope values also overlaps with other large sharks such as Araloselachus cuspidatus, an extinct sand tiger shark (Szabo & Kocsis 2016; Hovestadt 2020) that would’ve mainly eaten fish given its large, smooth, slender teeth (Cooper et al. 2023; McCormack et al. 2025). Secondly, the average zinc isotope values in Otodus species actually varied between different localities examined (Figure 2). Finally, a non-megalodon related result that stands out is that Carcharodon hastalis has significantly higher zinc isotope values than its living descendant, the modern great white shark (McCormack et al. 2025).
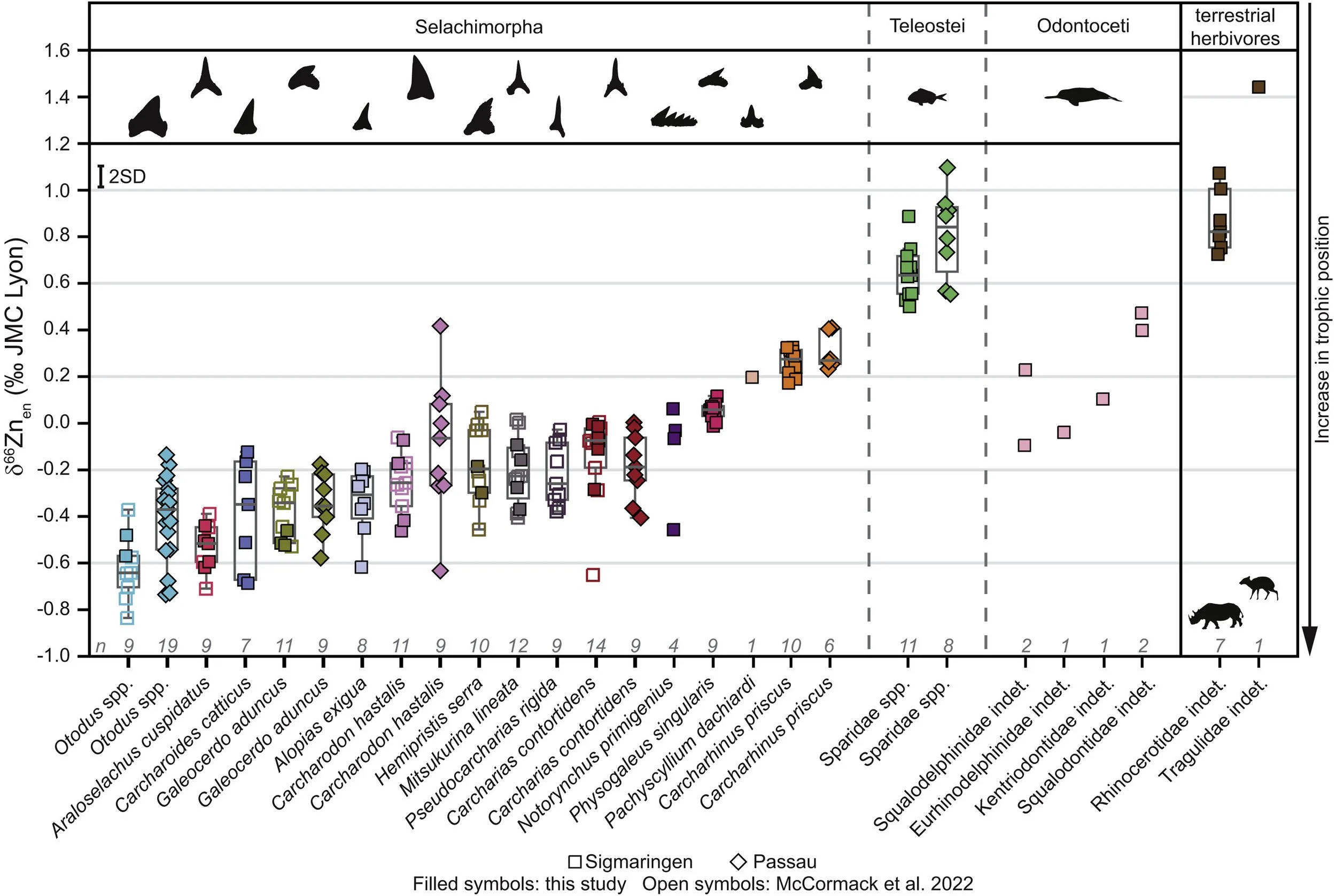
Figure 2. Zinc isotope values from the Early Miocene of Germany across two different formations; showcasing differences in trophic level between populations of the examined species. This figure is sourced from Figure 1 of McCormack et al. (2025).
What the study tells us about megalodon’s diet?
Okay, so what does all of this actually mean? It all comes down to lower zinc isotope values representing a higher trophic level. Otodus species having the very lowest zinc values clearly indicates that they did in fact occupy the very highest trophic levels (i.e., they were “supercarnivores”), as earlier work has already suggested (Martin et al. 2015; Kast et al. 2022; Cooper et al. 2022). Yet that range of values across individuals and localities also implies two caveats to this: (1) that they were also feeding on lower trophic levels and not just on species directly below them; and (2) their preferred diet likely differed across populations. When it comes to white sharks, the living white shark’s mean zinc isotope value was not significantly different from megalodon or O. chubutensis, implying some trophic overlap as suggested by other works (McCormack et al. 2022; Paredes-Aliaga & Herraiz 2024). By contrast, C. hastalis having significantly higher zinc isotope values indicates that it was actually at a lower trophic level that its eventual descendant, likely preferring to eat fish rather than marine mammals (Figure 3; McCormack et al. 2025).

Figure 3. Zinc isotope values of Miocene and living shark teeth showcasing differences across both populations and species. This figure is sourced from Figure 4 of McCormack et al. (2025).
The central takeaway is summarised very well in the paper’s title, which calls megalodon and O. chubutensis “opportunistic supercarnivores” – with “opportunistic” being the key word here. On the surface, this might not seem an entirely surprising result for somebody well-versed in shark ecology. Large, apex predatory sharks are opportunistic by nature, and will never turn down an easy chance at a meal (O’Shea et al. 2015). The same would certainly have been true of the gigantic megatoothed sharks like megalodon and O. chubutensis. In megalodon’s case, we’re talking about a shark that may have needed to eat the equivalent of over 90,000 kcal every single day to justify being so big (Cooper et al. 2022). So those of us who have worked on megalodon ecology would have expected it to be opportunistic and feed on whatever was available to it. What makes this new study (McCormack et al. 2025) so important is that it provides concrete data for this ecology, and also strongly indicates that megalodon was feeding on different kinds of prey across its different populations. Keep in mind that megalodon teeth are found all over the world (Pimiento et al. 2016; Pollerspöck et al. 2023) and so different habitats would have provided different prey preferences for different megalodon populations, which may have also varied in average size (Pimiento & Balk 2015; Shimada et al. 2022). Across its different populations, megalodon was not simply specialising in whales – although it very much was eating that prey – and instead may have primarily fed on smaller marine mammals or other prey items across its various habitats. For the fossil white sharks, the living great white shark didn’t just overlap in trophic level with at least some megalodon populations but were also higher-level predators than their ancestors.
What does this potentially mean for megalodon’s extinction in the Pliocene (Pimiento & Clements 2014; Boessnecker et al. 2019)? Well, it means that megalodon was certainly capable of exploiting the same resources as the living great white shark, specifically eating the same prey such as small marine mammals. This implies that some kind of competition may have taken place at the end of megalodon’s reign. A question I would like to see explored is if total length itself has a significant effect on isotopic values within megalodon and white sharks. The crown height-based equations used in the study give conservative values for megalodon (see Shimada et al. 2025) and, based on this, the size of the megalodons in this study ranged from 6-10 m in length (McCormack et al. 2025). While we don’t know megalodon’s different life stages, taking a maximum size of 20-24 m (Perez et al. 2021; Shimada et al. 2025) at face value would imply that these may well have been juveniles; and certainly a similar size to the very largest white sharks, which reach about 6 m in length (Randall 1973; De Maddalena et al. 2003; Christensen et al. 2014). Nevertheless, megalodon was clearly not just a specialist in eating big whales but probably ate whatever it wanted based on availability – be it a big fish, a small seal, another large shark, a baleen whale, or a carcass ripe for the picking (McCormack et al. 2025).
Isotope studies are a recent but highly welcome methodology in the quest to unravel megalodon’s dietary ecology. What at first appeared to be a story of two contrasts has now nicely come together to reveal that megalodon was a predator at the very top of the food web, beyond anything in our oceans today, and yet it was also opportunistic to whatever was unlucky enough to be in its way (Martin et al. 2015; Kast et al. 2022; McCormack et al. 2022; 2025). These sorts of studies have also been useful in other ways for megalodon. For example, the extraction of oxygen isotopes from teeth has allowed for body temperature estimates confirming the presence of regional endothermy in this species (Ferrón 2017; Griffiths et al. 2023). Whatever comes next for isotopic studies focusing on megalodon, I have no doubt they will reveal even more about the giant shark’s diet, and I await those results with anticipation.
Further reading
You can read the full paper at Earth and Planetary Science Letters under the following reference:
McCormack J, Feichtinger I, Fuller BT, Jaouen K, Griffiths ML, Bourgon N, Maisch IV H, Becker MA, Pollerspöck J, Hampe O, Rössner GE, Assemat A, Müller W & Shimada K, 2025. Miocene marine vertebrate trophic ecology reveals megatooth sharks as opportunistic supercarnivores. Earth and Planetary Science Letters, 119392.
References
Aguilera OA, García L & Cozzuol MA, 2008. Giant-toothed white sharks and cetacean trophic interaction from the Pliocene Caribbean Paraguaná Formation. Paläontologische Zeitschrift, 82, 204-208.
Assemat A, Adnet S, Bayez K, Hassler A, Arnaud‐Godet F, Mollen FH, Girard C & Martin JE, 2022. Exploring diet shifts and ecology in modern sharks using calcium isotopes and trace metal records of their teeth. Journal of Fish Biology, 105, 1469-1481.
Bird H, 2022. When ocean giants collide. Palaeontologia Electronica blog. https://palaeo-electronica.org/content/blog/3700-megalodon-whale-shear-compression-fracture.
Boessenecker RW, Ehret DJ, Long DJ, Churchill M, Martin E & Boessenecker SJ, 2019. The Early Pliocene extinction of the mega-toothed shark Otodus megalodon: a view from the eastern North Pacific. PeerJ, 7, e6088.
Casey MM & Post DM, 2011. The problem of isotopic baseline: reconstructing the diet and trophic position of fossil animals. Earth-Science Reviews, 106, 131-148.
Christiansen HM, Lin V, Tanaka S, Velikanov A, Mollet HF, Wintner SP, Fordham SV, Fisk AT & Hussey NE, 2014. The last frontier: catch records of white sharks (Carcharodon carcharias) in the Northwest Pacific Ocean. PLoS One, 9, e94407.
Collareta A, Lambert O, Landini W, Di Celma C, Malinverno E, Varas-Malca R, Urbina M & Bianucci G, 2017. Did the giant extinct shark Carcharocles megalodon target small prey? Bite marks on marine mammal remains from the late Miocene of Peru. Palaeogeography, Palaeoclimatology, Palaeoecology, 469, 84-91.
Cooper JA, Griffin JN, Kindlimann R & Pimiento C, 2023. Are shark teeth proxies for functional traits? A framework to infer ecology from the fossil record. Journal of Fish Biology, 103, 798-814.
Cooper JA, Hutchinson JR, Bernvi DC, Cliff G, Wilson RP, Dicken ML, Menzel J, Wroe S, Pirlo J & Pimiento C, 2022. The extinct shark Otodus megalodon was a transoceanic superpredator: Inferences from 3D modeling. Science Advances, 8, eabm9424.
Cooper JA, Pimiento C, Ferrón HG & Benton MJ, 2020. Body dimensions of the extinct giant shark Otodus megalodon: a 2D reconstruction. Scientific Reports, 10, 14596.
De Maddalena A, Glaizot O & Oliver G, 2003. On the great white shark, Carcharodon carcharias (Linnaeus, 1758), preserved in the Museum of Zoology in Lausanne. Marine Life, 13, 53-59.
Ehret DJ, MacFadden BJ, Jones DS, Devries TJ, Foster DA & Salas-Gismondi, R, 2012. Origin of the white shark Carcharodon (Lamniformes: Lamnidae) based on recalibration of the Upper Neogene Pisco Formation of Peru. Palaeontology, 55, 1139-1153.
Ferrón HG, 2017. Regional endothermy as a trigger for gigantism in some extinct macropredatory sharks. PLoS One, 12, e0185185.
Godfrey SJ & Altman J, 2005. A Miocene cetacean vertebra showing a partially healed compression fracture, the result of convulsions or failed predation by the Giant White Shark, Carcharodon megalodon. Jeffersoniana, 16, 1-12.
Godfrey SJ & Beatty BL, 2022. A Miocene cetacean vertebra showing a partially healed longitudinal shear-compression fracture, possibly the result of domoic acid toxicity or failed predation. Palaeontologia Electronica, 25, a28.
Godfrey SJ, Ellwood M, Groff S & Verdin MS, 2018. Carcharocles-bitten odontocete caudal vertebrae from the Coastal Eastern United States. Acta Palaeontologica Polonica, 63, 463-468.
Godfrey SJ, Nance JR & Riker NL, 2021. Otodus-bitten sperm whale tooth from the Neogene of the Coastal Eastern United States. Acta Palaeontologica Polonica, 66, 599-603.
Griffiths ML, Eagle RA, Kim SL, Flores RJ, Becker MA, Maisch IV HM, Trayler RB, Chan RL, McCormack J, Akhtar AA, Tripati AK & Shimada K, 2023. Endothermic physiology of extinct megatooth sharks. Proceedings of the National Academy of Sciences, 120, e2218153120.
Hovestadt D, 2020. Taxonomic adjustments of the Oligocene and Miocene Odontaspididae and Carchariidae based on extant specimens. Cainozoic Research, 20, 229-255.
Kallal RJ, Godfrey SJ & Ortner DJ, 2010. Bone reactions on a pliocene cetacean rib indicate short term survival of predation event. International Journal of Osteoarchaeology, 22, 253-260.
Kast ER, Griffiths ML, Kim SL, Rao ZC, Shimada K, Becker MA, Maisch HM, Eagle RA, Clarke CA, Neumann AN, Karnes ME, Lüdecke T, Leichliter JN, Martínez-García A, Akhtar AA, Wang XT, Haug GH & Sigman DM, 2022. Cenozoic megatooth sharks occupied extremely high trophic positions. Science Advances, 8, eabl6529.
Klimley AP, 1994. The predatory behavior of the white shark. American Scientist, 82, 122-133.
Martin RA, Hammerschlag N, Collier RS & Fallows C, 2005. Predatory behaviour of white sharks (Carcharodon carcharias) at Seal Island, South Africa. Journal of the Marine Biological Association of the United Kingdom, 85, 1121-1136.
Martin JE, Tacail T, Adnet S, Girard C & Balter V, 2015. Calcium isotopes reveal the trophic position of extant and fossil elasmobranchs. Chemical Geology, 415, 118-125.
McCormack J, Feichtinger I, Fuller BT, Jaouen K, Griffiths ML, Bourgon N, Maisch IV H, Becker MA, Pollerspöck J, Hampe O, Rössner GE, Assemat A, Müller W & Shimada K, 2025. Miocene marine vertebrate trophic ecology reveals megatooth sharks as opportunistic supercarnivores. Earth and Planetary Science Letters, 119392.
McCormack J, Griffiths ML, Kim SL, Shimada K, Karnes M, Maisch H, Pederzani S, Bourgon N, Jaouen K, Becker MA, Jöns N, Sisma-Ventura G, Straube N, Pollerspöck J, Hublin JJ, Eagle RA & Tütken T, 2022. Trophic position of Otodus megalodon and great white sharks through time revealed by zinc isotopes. Nature Communications, 13, 2980.
O’Shea OR, Mandelman J, Talwar B & Brooks EJ, 2015. Novel observations of an opportunistic predation event by four apex predatory sharks. Marine and Freshwater Behaviour and Physiology, 48, 374-380.
Paredes-Aliaga MV & Herraiz JL, 2024. Analysing trophic competition in †Otodus megalodon and Carcharodon carcharias through 2D-SEM dental microwear. Spanish Journal of Palaeontology, 39, 91-102.
Perez VJ, Leder RM & Badaut T, 2021. Body length estimation of Neogene macrophagous lamniform sharks (Carcharodon and Otodus) derived from associated fossil dentitions. Palaeontologia Electronica, 24, a09.
Pimiento C & Balk MA, 2015. Body-size trends of the extinct giant shark Carcharocles megalodon: a deep-time perspective on marine apex predators. Paleobiology, 41, 479-490.
Pimiento C & Clements CF, 2014. When did Carcharocles megalodon become extinct? A new analysis of the fossil record. PLoS One, 9, e111086.
Pimiento C, Kocáková K, Mathes GH, Argyriou T, Cadena EA, Cooper JA, Cortés D, Field DJ, Klug C, Scheyer TM, Valenzuela-Toro AM, Buess T, Günter M, Gardiner AM, Hatt P, Holdener G, Jacober G, Kobelt S, Masseraz S, Mehli I, Reiff S, Rigendinger E, Ruckstuhl M, Schneider S, Seige C, Senn N, Staccoli V, Baumann J, Flüeler L, Guevara LJ, Ickin E, Kissling KC, Rogenmoser J, Spitznagel D, Villafaña JA & Zanatta C, 2024. The extinct marine megafauna of the Phanerozoic. Cambridge Prisms: Extinction, 2, 1-17.
Pimiento C, MacFadden BJ, Clements CF, Varela S, Jaramillo C, Velez‐Juarbe J & Silliman BR, 2016. Geographical distribution patterns of Carcharocles megalodon over time reveal clues about extinction mechanisms. Journal of Biogeography, 43, 1645-1655.
Pollerspöck J, Cares D, Ebert DA, Kelley KA, Pockalny R, Robinson RS, Wagner D & Straube N, 2023. First in situ documentation of a fossil tooth of the megatooth shark Otodus (Megaselachus) megalodon from the deep sea in the Pacific Ocean. Historical Biology, 37, 120-125.
Randall JE, 1973. Size of the great white shark (Carcharodon). Science, 181, 169-170.
Shimada K, 2003. The relationship between the tooth size and total body length in the white shark. Journal of Fossil Research, 35, 28-33.
Shimada K, Maisch IV HM, Perez VJ, Becker MA & Griffiths ML, 2022. Revisiting body size trends and nursery areas of the Neogene megatooth shark, Otodus megalodon (Lamniformes: Otodontidae), reveals Bergmann’s rule possibly enhanced its gigantism in cooler waters. Historical Biology, 35, 208-217.
Shimada K, Motani R, Wood JJ, Sternes PC, Tomita T, Bazzi M, Collareta A, Gayford JH, Türtscher J, Jambura PL, Kriwet J, Vullo R, Long DJ, Summers AP, Maisey JG, Underwood C, Ward DJ, Maisch HM, Perez VJ, Feichtinger I, Naylor GJP, Moyer JK, Higham TE, da Silva JPCB, Bornatowski H, González-Barba G, Griffiths ML, Becker MA & Siversson M, 2025. Reassessment of the possible size, form, weight, cruising speed, and growth parameters of the extinct megatooth shark, Otodus megalodon (Lamniformes: Otodontidae), and new evolutionary insights into its gigantism, life history strategies, ecology, and extinction. Palaeontologia Electronica, 28, a12.
Szabó M & Kocsis L, 2016. A new Middle Miocene selachian assemblage (Chondrichthyes, Elasmobranchii) from the Central Paratethys (Nyirád, Hungary): implications for temporal turnover and biogeography. Geologica Carpathica, 67, 573-594.
Wroe S, Huber DR, Lowry M, McHenry C, Moreno K, Clausen P, Ferrara TL, Cunningham E, Dean MN & Summers AP, 2008. Three‐dimensional computer analysis of white shark jaw mechanics: how hard can a great white bite? Journal of Zoology, 276, 336-342.